Polyploidization (whole-genome duplication) is among the most important evolutionary forces driving the diversification of flowering plants. By providing an almost instantaneous reproductive isolation of polyploid mutants from diploid progenitors, polyploidization has been widely acknowledged as the most frequent mode of sympatric speciation. Additionally, polyploidization brings genetic and phenotypic novelty, some of which is already apparent in first-generation polyploid mutants, though it remains unclear whether the new traits may increase their adaptive potential or facilitate their establishment in diploid progenitors’ populations. Polyploids tend to originate recurrently and many plant species retain individuals of two or more different ploidy levels in certain parts of their distributional range of even within their populations.
In our research, we aim to elucidate the key evolutionary mechanisms operating at various stages of polyploid speciation, which are important for the potential evolutionary success of polyploid lineages:
Rates of polyploid origin in natural populations and their environmental and genetic determinants
The dominant pathways of polyploid formation involve gametes with somatic chromosome number (i.e., unreduced gametes, 2n gametes) that occur in natural populations because of meiotic errors. First-generation tetraploid mutants may either directly result from a fusion of two unreduced gametes of diploids, or a single unreduced gamete may produce a triploid intermediate that can give rise to tetraploid offspring in crosses with diploids and/or other triploids (i.e., the triploid bridge mechanism). Generally, the rates of gametic non-reduction are very low (< 1% of produced gametes), though increased rates may be observed if plants are experiencing environmental stress, and natural populations may also contain individuals genetically predisposed for (heritable) high 2n-gamete production.
We are interested in the patterns of 2n gamete production in natural populations occurring along environmental gradients, in the relative importance of genetic and environmental factors (incl. the genotype × environment interaction), and in genetic architecture of 2n gamete production (and whether it can be generalised across populations and species).
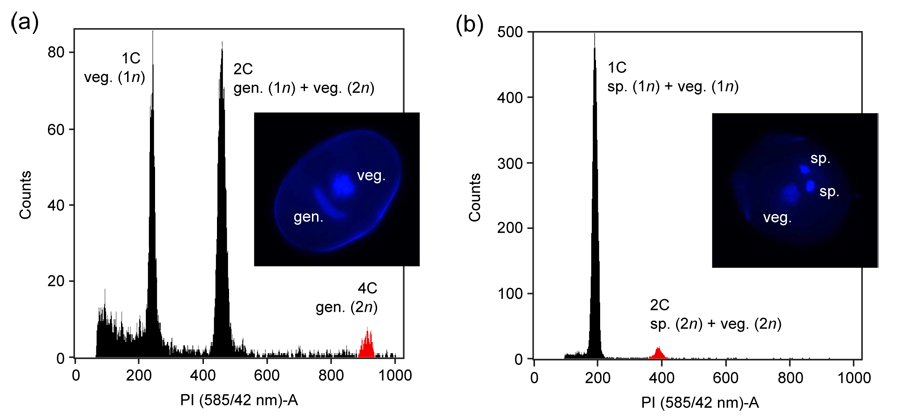
To infer the rates of 2n gametes we are using flow cytometry (FCM) of field-collected pollen samples. This technique allows rapid and cost-effective simultaneous analysis of thousands of nuclei per flower and thus reaching robust estimates of 2n gamete frequency. An overview of the currently used protocols and some best practices guidelines for pollen FCM can be found in this review article prepared in collaboration with the Husband lab (University of Guelph, Canada) and the FLOWer lab (University of Coimbra, Portugal).
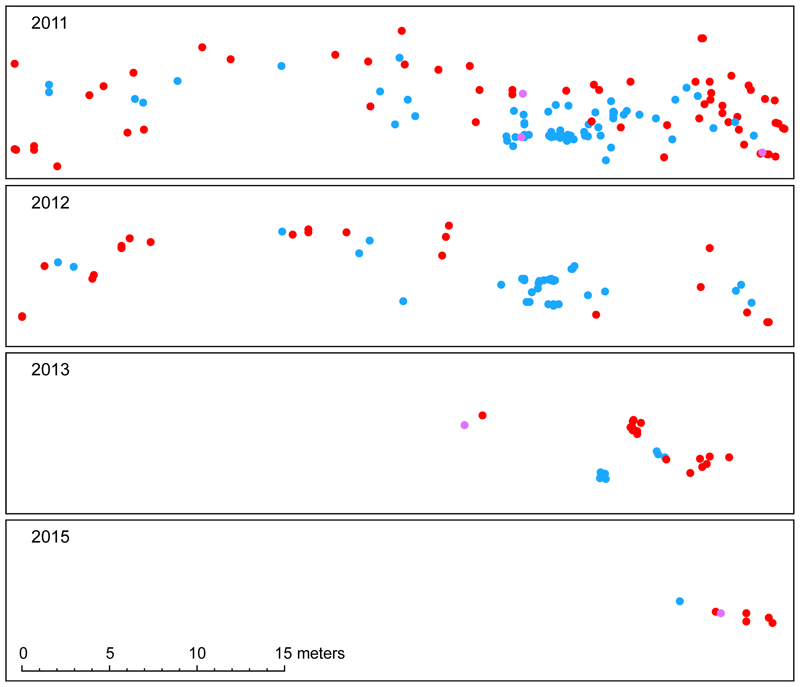
Obtaining direct estimates of the actual rate with which new polyploid mutants emerge in natural populations is, on the other hand, quite challenging. The early-generation polyploid mutants first need to be distinguished from their long-established polyploid counterparts (and inter-ploidy hybrids) and application of molecular markers is usually not compatible with the sample size required. In the scentless mayweed (Tripleurospermum inodorum), one of our studied mixed-ploidy species, we leveraged the existing 19% difference in monoploid genome size between diploids and tetraploids (enabling the easy distinction 3x and 4x mutants solely based on their nuclear DNA content in routine FCM ploidy screening) to provide the first robust estimates of the rate of spontaneous polyploidization in a plant species. Polyploid mutants were extremely rare among the 11,018 individuals screened across 1,209 T. inodorum populations and accounted for 0.04% of samples. They never formed uniform populations but occurred solitarily, without any sign of their local establishment and further spread (original article).
Conditions favouring local establishment of polyploid mutants in diploid progenitors’ populations
While the probability of spontaneous polyploidization in natural populations should increase under higher rates of 2n gamete production, new polyploid mutants are facing a strong demographic disadvantage that usually leads to their extirpation in progenitors’ populations. …
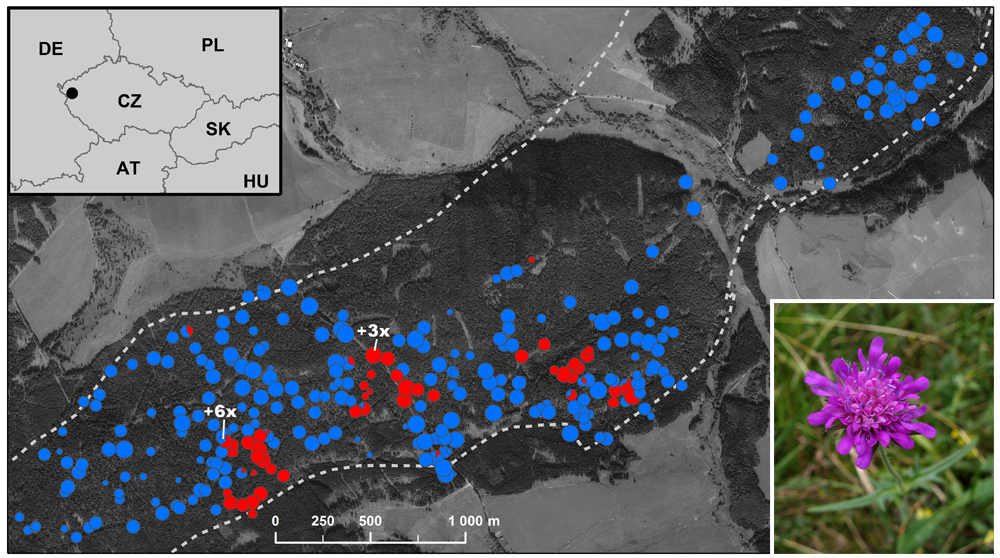

Phenotypic novelty originating as a direct consequence of whole-genome duplication and its putative adaptive potential
Text …
(In-)stability of diploid – polyploid coexistence within populations and contact zones
Text …