
1. Úvod
Parazitický prvok Toxoplasma gondii je jedním z nejrozšířenějších parazitů člověka. Odhaduje se, že v současnosti je infikováno 30-60% celkové lidské populace (Remington a spol. 1992 dle Holliman 1997). Toxoplasmosa, onemocnění jež tento parazit způsobuje, byla středem zájmu mnoha studií. Primát v tomto ohledu drží i česká věda. Poprvé byla toxoplasmosa u člověka zaznamenána v Praze roku 1923 (Janků 1923). V současnosti jsou celkem dobře známy klinické následky kongenitální i akutní formy toxoplasmosy. Nicméně nejrozšířenější formou toxoplasmosy je latentní onemocnění. Překvapivě málo pozornosti bylo věnováno této formě infekce. Obvykle je latentní toxoplasmosa považována za asymptomatickou s minimem klinických následků. Ukazuje se však, že spíše než klinické následky má latentní toxoplasmosa dopad na chování člověka (Flegr a spol. 1996).
V poslední době byly u mnoha různých živočichů zjištěny změny v chování způsobené parazity (Moore, Gotteli 1990). Často jsou změny v chování hostitele považovány za manipulaci ze strany parazita. Účelem manipulační aktivity parazita je zvýšení pravděpodobnosti přenosu do dalšího hostitele. V mnoha případech jsou však změny v chování parazitovaných jedinců buď pouze vedlejším produktem onemocnění a nebo adaptací hostitele na negativní působení parazita. Protože evoluce parazita a hostitele neprobíhala odděleně, ale ve vzájemných interakcích, je velmi obtížné rozhodnout, co je příčinou pozorovaných změn v chování. Přestože chování parazitovaných zvířat bylo objektem zájmu mnoha studií, poměrně malé množství prací se zabývalo behaviorálními následky parazitárních infekcí u člověka.
Tato práce se zabývá následky latentní toxoplasmosy na lidský organismus. Pozornost byla zaměřena nejen na zdravotní následky infekce, ale i na behaviorální změny způsobené latentní toxoplasmosou. Práce vznikla ve spolupráci kateder antropologie a parazitologie. Přestože se nejedná o spojení příliš obvyklé, věřím, že překračování hranic vlastních oborů bylo v tomto případě přínosné.
2. Literární přehled
2.1. Toxoplasma gondii
2.1.1.Systematické zařazení
Intracelulární parazitický prvok Toxoplasma gondii (Nicolle et Manceau) byl objeven roku 1908 u hlodavce gondiho saharského, Ctenodactylus gundi, od něhož je odvozeno druhové jméno parazita. Sami objevitelé však řadili tohoto parazita mezi původce leischmaniosy. Komplikace v systematickém zařazení organismu způsoboval i neznámý životní cyklus. Ten popsal až roku 1965 Hutchison. V současnosti je prvok Toxoplasma gondii řazen do čeledi Sarcocystidea, řádu Eimeriida, třídy Coccidea, kmene Apicomplexa (Levine 1992).
2.1.2. Životní cyklus prvoka Toxoplasma gondii
Životní cyklus toxoplasmy může být jak monoxenní tak i dixenní. V prvním případě se parazit přenáší mezi kočkovitými šelmami (čeleď Felidae). Přenos může probíhat jednak horizontálně, to znamená nakažením kočky oocystami vyloučenými jinou kočkou (Dubey a spol. 1970a), tak i vertikálně čili z matky na plod . V případě dixenního životního cyklu se toxoplasma přenáší do konečného hostitele - kočkovité šelmy pozřením nakaženého mezihostitele (Dubey a spol. 1970b), jímž může být zřejmě jakýkoliv homoiotermní živočich (Hutchison & Work 1969). Mimo to se může parazit šířit mezi intermediálními hostiteli a to opět dvojím způsobem horizontálně - predací a vertikálně- kongenitálním přenosem z matky na plod (Owen & Trees 1998).
Parazit je schopen infikovat buňky nejrůznějších tkání hostitele. Nejčastěji však napadá tkáně svalové a mozkové. Rozmnožování probíhá vždy intracelulárně. K vytváření gamet a pohlavnímu rozmnožování dochází ve střevním epitelu kočky, především v ileální části tenkého střeva. Gametogonii předchází množství generací nepohlavního množení prostředenictvím zvláštního typu schizogonie. Při tomto typu dělení dochází k vícenásobnému rozdělení jader bez rozdělení cytoplasmy a k vytvoření dceřinných buněk uvnitř buňky mateřské (Hutchison a spol. 1971).
Toxoplasma gondii může existovat ve třech různých životních formách: sporozoiti, tachyzoiti a bradyzoiti. Jednotlivé životní formy se liší svým výskytem v jednotlivých fázích nákazy i infekčností při přenosu do dalšího hostitele. Ultrastrukturální stavba buněk jednotlivých životních forem je velmi podobná, rozdíly jsou nacházeny ve stavbě rhoptrií (struktury exkrečního aparátu na apikálním konci buňky) a v množství densních granulí (Dubey a spol. 1998).
Sporozoiti jsou stadium vznikající jako produkt pohlavního rozmnožování ve střevě kočky. Přímo po oplození vzniká zygota, z níž se formují dvě sporocysty. Postupnou sporulací se vytvářejí v každé sporocystě čtyři sporozoiti a celý útvar se nazývá oocysta (Dubey a spol. 1970a). Oocysty jsou vylučovány nevysporulované v kočičím trusu. K vylučování oocyst dochází po několika dnech od nákazy po dobu okolo jednoho týdne (Hagiwara a spol.1981; Dubey a spol. 1970b). V začátku vylučování i jeho délce jsou značné rozdíly závislé především na životní formě parazita, jímž se kočka infikuje. Po pozření oocysty hostitelem dochází v prostředí žaludečních šťáv k excystaci a uvolnění sporozoitů. Sporozoiti pronikají do tkání hostitele a mění se na tachyzoity.
Jako tachyzoiti jsou označována stadia rychle se množící v jakýchkoliv tkáních mezihostitele a mimo střevní epitel konečného hostitele. (Dubey a spol. 1998) Rozmnožování tachyzoitů probíhá specializovanou formou asexuální reprodukce tzv. endodyogenií. Ta je charakterizována vznikem dvou dceřinných buněk uvnitř buňky mateřské.
Bradyzoiti jsou pomalu se množícím stadiem uvnitř tkáňových cyst. Tvorba tkáňových cyst je zřejmě reakcí parazita na imunitní odpověď hostitele (Gross a spol. 1996). Tkáňové cysty jsou schopny perzistovat v hostiteli po celý jeho život (Dubey 1986). Mohou se vyskytovat v kterékoliv tkáni, nejčastěji jsou nacházeny v tkáních nervových a svalových. V některých případech např. po imunosupresi nebo u pacientů s AIDS, může dojít k opětné konverzi bradyzoitů na tachyzoity. Bradyzoiti jsou oproti tachyzoitům odolnější proti působení gastrických šťáv(Sharma & Dubey1981). Přestože se kočky mohou nakazit všemi infekčními stadii, jejich infektivita se značně liší. Zatímco infektivita tachyzoitů a sporozoitů je nižší než 30%, po pozření bradyzoitů vylučovaly kočky oocysty téměř ve všech případech (Dubey & Frenkel 1976).
Nízká tkáňová a hostitelská specifita spolu s různými možnostmi přenosu jsou zřejmě rozhodujícími faktory z hlediska globálního rozšíření prvoka Toxoplasma gondii.
2.2.Toxoplasmosa
Onemocnění způsobené parazitem Toxoplasma gondii se nazývá toxoplasmosa. U člověka toto onemocnění poprvé posal roku 1923 Janků (Janků 1923). Podle způsobu nákazy se rozlišují dva základní typy onemocnění: toxoplasmosa kongenitální a získaná.
2.2.1.Kongenitální toxoplasmosa
K přenosu parazita na plod může u člověka dojít jen v případě akutní infekce matky v období těhotenství. Pokud byla žena nakažena již před otěhotněním, k přenosu na plod nedochází. Ženám nakaženým v období těhotenství se asi v 40% případů narodí dítě kongenitálně infikované. Zatímco pravděpodobnost přenosu parazita na dítě od prvního k třetímu trimestru stoupá, závažnost následků nákazy v průběhu těhotenství klesá. Nejvíce asymptomatických dětí se nakazí v posledním trimestru (Koppe & Rothova 1989). Nejčastějším následkem kongenitální toxoplasmosy je chorioretinitis a dále též hydrocefalus, intracerebrální kalcifikace a mikropthalmus. Často se též objevují záchvaty, motorické abnormálie a snížené IQ (Roizen a spol. 1995). Některé následky kongenitální toxoplasmosy především chorioretinitis se mohou projevit až po několika letech (Koppe & Rothova 1989; Pariser a spol. 1978).
2.2.2. Získaná toxoplasmosa
Postnatální infekce prvokem Toxoplasma gondii probíhá ve velkém množství případů asymptomaticky. Nejčastějším příznakem akutní fáze infekce je lymphadenopatie zejména v oblasti krku (Braveny a spol. 1973) a to i bez dalších symptomů. Souběžně se mohou objevit nespecifické příznaky jako je únava, malátnost, myalgie, bolesti hlavy a subfebrilní teplota, podobně jako u běžných virových onemocnění (Remington 1974; Černý a spol.1995). Tyto příznaky ve většině případů během několika týdnů spontánně vymizí. Chronická fáze infekce je pak obvykle latentní a asymptomatická. Jsou však popsány i případy chronické aktivní lymphoadenopatické formy toxoplasmosy u imunokompetentních pacientů (O’Connell a spol. 1993). K reaktivaci latentní nákazy může dojít při oslabení organismu jiným interkurentním onemocněním (např. tuberkulosa) (Petrovický & Vojtěchovský 1955), podáváním imunosupresiv (např. kortikostereoidy) (Frenkel a spol. 1975a) nebo u poruch imunitního systému (např. Hodgkinsova nemoc, AIDS) (Remington 1974).
V některých případech však dochází k postižení CNS, projevující se silnými bolestmi hlavy, intrakraniálním tlakem, epileptickými záchvaty, spavostí, depresemi a apatií. Často je pak diagnostifikována encephalitis, meningo-encephalitis či meningo-enephalo-myelitis toxoplasmového původu (Kramer 1966; Ladee a spol.1966; Petrovický & Vojtěchovský 1955). U některých pacientů dochází i k psychiatrickým poruchám: halucinacím, vzniku organické psychosy a progresivní demenci (Ladee a spol.1966). Chorioretintis se u získané, narozdíl od kongenitální, toxoplasmosy vyskytuje velmi zřídka (Koppe, Rothova 1989). Jsou známy i případy toxoplasmové myokarditis (Kouba a spol.1966), myositis a pneumonie (Jírovec 1977). Ve valné většině případů se však tyto symptomy nevyskytují izolovaně, ale jsou součástí tzv. generalizované formy toxoplasmosy, při níž může být napaden v podstatě kterýkoliv orgán a není -li léčena, končí obvykle fatálně (Remington 1974). Touto formou onemocnění jsou postihováni téměř výhradně imunodeficientní jedinci, tedy v součastnosti především pacienti s AIDS.
Důležitým markerem fáze infekce jsou hladiny jednotlivých typů protilátek. V akutní fázi infekce dochází k strmému vzestupu hladiny protilátek typu IgM , IgA a k pozvolnému vzestupu protilátek typu IgG. Pro chronickou fázi infekce je charakteristická přítomnost protilátek typu IgG, zatímco protilátky typu IgA a IgM obvykle vymizí do devíti měsíců od nákazy (Kodym, Tolarová 1998).
2.2.3. Epidemiologie toxoplasmosy
2.2.3.1. Prevalence toxoplasmosy
Parazit Toxoplasma gondii je kosmopolitně rozšířen a patří u člověka mezi nejrozšířenější infekční agens (Remington 1974). Přímý průkaz parazita je u živých jedinců velmi těžko uskutečnitelný, používá se proto k diagnostice toxoplasmosy nepřímých, imunologických metod. Ty jsou založeny na speciálních serologických technikách zjišťujících existenci specifických protilátek nebo na intradermálním testu zjišťujícím specifickou buněčnou imunitu. Na těchto testech je založena nejen klinická diagnostika, ale používají ji i všechny prevalenční studie. Přestože je toxoplasma v podstatě celosvětově rozšířena existují značné rozdíly v jejím výskytu mezi jednotlivými populacemi. V Evropě byla zjištěna velmi vysoká prevalence toxoplasmosy u těhotných matek v Paříži, 86% (Desmonts & Couvreur 1974), v jiných zemích je průměrná prevalence např. 31,8% v Rumunsku (Elias 1966), 38,8% v Maďarsku (Jankó & Czeizel 1970), 67% ve Španělsku (Perea & Borobio 1974), 45,2% v Hamburku (Schassan & Kaskara 1971). Nejnižší promořenost v Evropě byla zjištěna mezi Laponci ze severní Skandinávie 2,3% (Huldt a spol. 1979).
Studie z Afriky naznačují obdobnou prevalenci jako v Evropě např. 52,6% v Mali (Quilici a spol. 1976), 25,9% v Nigerii (Bozděch & Moronfoye 1974), 51,4% v Togu (Saathoff & Dogba 1974), 18,2% v Nigeru (Develoux a spol. 1988), 49,5% v Ghaně (Godwin & Remington 1973).
Velmi vysoká průměrná seroprevalence je v Střední a Jižní Americe např. 61% v Brazílii (Ricciardi a spol. 1978), 61,4% v Kostarice (Frenkel & Ruiz 1980), 38% v Peru (Cantella a spol. 1974), 26,2% v Mexiku (Resano a spol. 1985), 33,4% na Kubě (Garcia 1983), 59% v Salvádoru (Remington a spol. 1970).
Obecně nižší průměrná seroprevalence je v Asii např. 15,4% v Japonsku (Komiya a spol. 1961), 32,4% v Malaysii (Vijayamma a spol. 1980), 9,8% v Hong Kongu (Ko a spol. 1980).
V České republice byla na základě vyšetření 3342 sér zjištěna průměrná seroprevalence 35,5% (31% mužů a 39,9% žen) (Kodym a spol.1995). Seroprevalence v jednotlivých věkových skupinách je uvedena v tab. č 1.
|
věková kategorie (roky) |
pozitivita (%) |
||
|
muži |
ženy |
celkem |
|
|
0 - 4 |
13,7 |
26,9 |
20,3 |
|
5 - 9 |
26,1 |
29,4 |
27,7 |
|
10 - 14 |
21,6 |
33,7 |
27,7 |
|
15 - 19 |
28,3 |
39,2 |
33,8 |
|
20 - 29 |
36,6 |
40,2 |
38,4 |
|
30 - 39 |
28,4 |
44,9 |
36,7 |
|
40 - 49 |
35,6 |
46,9 |
41,3 |
|
50 - 59 |
34,4 |
41,9 |
38,2 |
|
60+ |
33,2 |
35,4 |
34,3 |
|
celkem |
31,0 |
39,9 |
35,5 |
Tab. č.1: Seroprevalence toxoplasmosy v České republice v jednotlivých věkových kategoriích. Podle Kodym a spol. (1995).
Prevalence toxoplasmosy narůstá s věkem (Konishi & Takahashi 1987; Bozděch & Moronfoye 1974; Ricciardi a spol. 1978; Barbier a spol. 1983). V některých zemích např. v Salvádoru (Remington a spol. 1970) dosahuje prevalence toxoplasmosy ve věkových skupinách nad 50 let více jak 90%. Kumulativní nárůst prevalence je dán doživotní perzistencí parazita v tkáních hostitele. V některých studiích byl však zjištěn pokles seroprevalence ve věkových skupinách nad 50let (Hejlíček a spol. 1993; De Roever-Bonnet 1967). Vysvětlení můžeme hledat ve vyhasínající odpovědi na infekci (Kodym a spol. 1995). Může jít také o artefakt průřezových studií v nichž se odráží i epidemiologická situace minulých let.
Statisticky signifikantní sexuální rozdíly v prevalenci nebyly ve většině studií zjištěny (Johnson a spol. 1980; Gandahusada 1978; Cantella a spol. 1974; Ricciardi a spol. 1978; Monjour a spol. 1983; De Roever-Bonnet 1972). Některé práce však uvádějí vyšší prevalenci u žen (Huldt a spol. 1979; Perea & Borobio 1974) jiné vyšší u mužů (Konishi & Takahashi 1987; Bozděch & Moronfoye 1974). Rozdíly v sociálních rolích mužů a žen v některých kulturách, a z nich vyplývající rozdíly v chování, mají zřejmě vliv i na pravděpodobnost nákazy parazitem Toxoplasma gondi. Odrazem specifických mezipohlavních rozdílů v rolích u některých populacích jsou pak i mezipohlavní rozdíly v prevalenci toxoplasmosy.
2.2.3.2. Rizikové faktory
Člověk se může postnatálně nakazit toxoplasmosou pozřením masa obsahujícím tkáňové cysty nebo materiálem kontaminovaným oocystami vylučovaných v kočičím trusu. Jedním z rozhodujících faktorů uplatňujících se v epidemiologii toxoplasmosy jsou tedy potravní zvyky. Konzumace syrového nebo špatně tepelně upraveného masa zvyšuje pravděpodobnost nakažení se toxoplasmosou (Braveny a spol. 1973; Stagno & Thiermann 1973; Buffolano a spol. 1996). Byly popsány i případy infekce toxoplasmosy přímo po požití syrového masa (Sacks a spol. 1983). Vysoká prevalence toxoplasmosy je v některých zemích např. ve Francii vysvětlována oblibou jídel z ne zcela tepelně upraveného masa (Desmonts & Couvreur 1974) a naopak nízká prevalence nepřítomností těchto praktik např. v čínských komunitách v Hong Kongu (Ko a spol. 1980) i v Jakartě (Gandahusada 1978). Výše rizika při konzumaci syrového masa samozřejmě závisí také na druhu zvířete z něhož maso pochází. Nejrizikovější je v tomto ohledu králičí maso z malochovů u něhož byla v České republice zjištěna promořenost 53% (Hejlíček & Literák 1994) a maso skopové u něhož byla promořenost toxoplasmosou 54,6% (Hejlíček & Literák 1993a). Podstatně nižší prevalence toxoplasmosy byla zjištěna u jatečných prasat 5,9% (ovšem 35,1% v domácích chovech) (Hejlíček, Literák 1993b) a podobně nízká prevalence u skotu 4,1% (Hejlíček & Literák 1992). Studie z Panamy (Frenkel a spol.1995),La Guadaloupe (Barbier a spol. 1983) a Švédska (Huldt a spol. 1979) vztah mezi konzumací syrového masa a toxoplasmosou nezjistily.
Druhým zdrojem infekce jsou oocysty vylučované v kočičím trusu. Výsledky prací zjišťujících asociaci mezi kontaktem s kočkou a toxoplasmosou se různí. Zatímco studie z Ruska (Ševkunova a spol. 1976), Kostariky (Frenkel & Ruiz 1980) a La Guadaloupe (Barbier a spol. 1983) tento vztah zjistily, ve studiích z Japonska (Konishi & Takahashi 1987), USA (DiGiacomo a spol. 1990), Německa (Braveny a spol. 1973) a Itálie (Buffolano a spol. 1996) vztah mezi kontaktem s kočkou a toxoplasmosou zjištěn nebyl. Mezi původními obyvateli na Nové Guiney byly v oblastech, v nichž jsou domácí kočky chovány zcela vyjímečně nebo vůbec, detekovány protilátky proti Toxoplasma gondii v méně jak 2% případů narozdíl od 14% do 34% pozitivně reagujících případů v oblastech kde byl chov koček běžnější (Wallace a spol. 1974). Pokud je kočka chována izolovaně v bytě a krmena konzervovaným masem je riziko její nákazy, a tím pádem i možnost nákazy člověka, velmi malé (Frenkel & Dubey 1972). Protože oocysty vyloučené v kočičím trusu jsou infekční až po vysporulování tzn. po 2-4 dnech, je doporučováno denně čistit nádobu, do níž kočky defekují (Robinson 1973). Kočka vylučuje oocysty po několika dnech od nakažení po dobu asi sedmi dnů (Hagiwara a spol. 1981). Přestože byly zjištěny případy reaktivace vylučování oocyst, většina koček vylučuje pouze jednou za život (Robinson 1973).
Zvýšené riziko nakažení se Toxoplasma gondii neplatí pouze pro kontakt s kočkou jako přímým zdrojem infekce, ale i pro kontakt s materiály kontaminovanými infikovanými výkaly. Velký význam mají v tomto ohledu podrobné studie Frenkela a Ruize z Kostariky (Frenkel & Ruiz 1980; Frenkel & Ruiz 1981). Podle autorů se lidé mající kočku často nakazí uvnitř domu, především v domech s betonovou podlahou, kde kočky defekují. Vysoká prevalence je i u lidí žijících v domech s dřevěnou podlahou, v nichž bývají často prostory sloužící volně žijícím kočkám jako úkryt a jsou tak zdrojem kontaminované půdy. Oocysty jsou schopny v prostředí dlouhodobě přežívat. V zemi zůstávaly infekční i po osmnácti měsících (Frenkel a spol. 1975b). Délka přežívání oocyst závisí zejména na teplotě a vlhkosti. V suchém prostředí a při vystavení přímému slunečnímu světlu dochází k jejich rychlé destrukci (Yilmaz & Hopkins 1972). Na ostrově La Guadeloupe byla v oblastech s větším množstvím srážek vyšší prevalence toxoplasmosy než v sušších oblastech (Barbier a spol. 1983), studie ze Salvádoru však tuto asociaci nezjistila (Remington a spol. 1970). V Peru byla zjištěna negativní korelace mezi prevalencí toxoplasmosy a nadmořskou výškou (Cantella a spol. 1974).
Rizikovými profesemi z hlediska toxoplasmosy jsou ty, při nichž dochází k manipulaci s infikovaným či kontaminovaným materiálem. Takovou profesí je např. práce na jatkách (Fayomi a spol. 1987; Nemec a spol. 1988; Elias 1966).
Vyšší prevalence toxoplasmosy u lidí žijících na venkově oproti obyvatelům měst je vysvětlována častějším kontaktem s půdou, která může být případně kontaminována oocystami (Schnurrenberger a spol. 1964; Jíra & Rosický 1983). Rozdíly v prevalenci toxoplasmosy podle místa bydliště však jiní autoři nenalezli (Gibson 1956).
Důležitou roli v mechanickém přenosu Toxoplasma gondii mohou hrát psi, u nichž je častá koprofílie (Frenkel a spol. 1995).
Rozdílné výsledky jednotlivých epidemiologických studií mohou být odrazem specifické epidemiologické situace v jednotlivých zemí. Zatímco např. v Evropě hrají v epidemiologii toxoplasmosy významnou roli potravní zvyky - tedy přenos tkáňovými cystami, např. v zemích Střední Ameriky jsou zřejmě rozhodující faktory spojené s přenosem oocystami.
2.3. Změny v chování hostitele následkem parazitace
Chování parazitovaných jedinců se často liší od těch, kteří parazitováni nejsou. U subhumánních organismů byla zjištěna celá řada typů chování, které může být v důsledku parazitace pozměněno.
Nejčastěji dochází ke změnám v aktivitě. Někteří parazité způsobují snížení aktivity hostitele, naopak infekce jinými parazity je spojena se zvýšením aktivity hostitele. Např. myši nakažené svalovcem Trichinella spiralis vykazovaly signifikantně nižší aktivitu v open field testu oproti kontrolám (Rau 1983). Naopak infekce škrkavkou Toxocara canis způsobovala u myší hyperaktivitu (Hay a spol. 1986).
Parazit také může měnit chování v neznámém prostředí nebo reakci hostitele na nový stimul. Např. u myší infikovaných krevničkou Schistosoma mansoni došlo k redukci exploračního chování (panáčkování) na polovinu v porovnání s kontrolami (Fiore a spol. 1996).
Následkem parazitace může též dojít ke změnám ve schopnosti učení a paměťových funkcí. Např. infekce helmintem Heligmosomoides polygyrus způsobuje u myší zhoršení prostorového učení (testováno v tzv. vodním bludišti) (Kavaliers & Colwell 1995a).
Parazitovaní jedinci mají také často pozměněné potravní nároky. Takže může docházet i ke změnám v potravním chování. Parazité používající k přenosu jako vektor hematofágní hmyz, často mění potravní chování vektora. Např. komáři Anopheles spp. infikovaní Plasmodium spp. sají signifikantně častěji oproti neinfikovaným komárům, čímž zvyšují pravděpodobnost transmise parazita (Morell 1997).
Důsledkem parazitace mohou být i změny v sociálním a sexuálním chování. Např. infekce svalovcem Trichinella spiralis způsobovala u myší snížení frekvence sociálních interakcí (Edwards 1988). V oblasti sexuálního chování bylo např. zjištěno, že samice myší dávají přednost pachu samcům nenakaženým oproti samcům nakaženým kokcidií Eimeria vermiformis (Kavaliers & Colwell 1995b).
V současné době byla behaviorálním následkům parazitace u zvířat věnována celá řada studií. Naproti tomu u člověka je vliv parazitů na chování, pomineme-li přímé následky onemocnění, jež jsou objektem zájmu klinických oborů, poměrně málo prozkoumanou oblastí. Bylo zjištěno, že infekce některými parazity je u člověka spojena se změnami v aktivitě, kognitivních schopnostech a dokonce i se změnami v osobnosti. V dalším textu (kap. 3.2. až 3.4.) jsou shrnuty současné poznatky o změnách v chování člověka způsobených parazity.
2.3.1. Příčiny změn
Jak bylo naznačeno na příkladu laboratorní myši u parazitovaných jedinců může docházet ke změnám v různých oblastech chování. Otázkou však je, co je příčinou pozorovaných změn. Nalézání odpovědi je často velmi obtížné, protože výsledný efekt je vždy výsledkem interakce mezi parazitem a jeho hostitelem. V zásadě mohou být změny v chování buď vedlejším produktem onemocnění, adaptací hostitele a nebo se též může jednat o adaptaci parazita.
2.3.1.1. Vedlejší produkt onemocnění
Tradičně jsou změny v chování spojovány s přímým patologickým působením parazita. Behaviorální následky patologického působení závisí samozřejmě především na druhu parazita a hostitele. Obecně je však možno říci, že parazité zvyšují energetickou náročnost hostitele, ať už je to spojeno s reparací parazitem poškozených tkání, přímou výživou parazita, zhoršením asimlační účinnosti, imunitní odpovědi či s dalšími mechanismy. Nedostatek energie se projeví jako snížení aktivity, letargie a deprese, což jsou obvyklé symptomy aktivního onemocnění (Holmes & Zohar 1990).
2.3.1.2. Adaptace hostitele
V evolučním boji s parazity se u hostitele vyvinulo množství obranných mechanismů (např. imunitní systém). Mezi takové obranné mechanismy patří i vyhýbání se parazitům čili jedná se o takové chování, které bude snižovat pravděpodobnost infekce. Například samice myši dokážou rozlišit mezi pachem parazitovaného a neparazitovaného samce a k pachu parazitovaných samců mají averzi (Kavaliers & Colwell 1995a; Kavaliers & Colwell 1995b). U člověka je vyhýbání se parazitům často spojeno s nejrůznějšími kulturními zákazy (tabu). Některá náboženství (židovské, islám) zakazují požívání vepřového masa, které má zásadní význam pro přenos některých helmintů (Trichinella spiralis, Ascaris lumbricoides). Korán také zakazuje defekovat a močit v místech zdrojů pitné vody a vody používané ke koupání. Restrikce takového chování má mimo jiné význam pro šíření původců schistosomiasy (Nelson 1990).
Některé změny v chování parazitovaného organismu jsou adaptacemi hostitele vedoucí k eliminaci parazita či kompenzaci jeho patologického působení. Sem je možno zařadit i u živočichů známou sebemedikaci, čili používání látek, většinou rostlinného původu, které mají negativní dopad na parazita a mnohdy vedou i k jeho eliminaci (Poulin 1995). V případě infekce ektoparazity se mohou změny v chování hostitele týkat množství času věnovaného čištění za účelem odstranění parazitů. Adaptací by mohla být za určitých podmínek i sebevražda hostitele, čímž by zabil nejen sebe, ale i parazita a tím by mohl uchránit od parazitace své příbuzné a zvýšit tak svoji inkluzivní fitnes (Smith Trail 1980).
Pokud není hostitel schopen parazita eliminovat, může změnit své chování tak, aby byl alespoň minimalizován dopad parazita na fitnes hostitele (Poulin 1995). Větší energetická náročnost parazitovaného jedince se tak může například projevit zvýšením času stráveného sháněním potravy. Příkladem kompenzace vlivu parazita je studie Parkerové (1992). Autorka studovala dopad schistosomiasy na denní aktivitu žen ze súdánského venkova. Nakažené ženy sbíraly stejné množství bavlny jako ženy nenakažené. Strávily však sběrem více času, čímž kompenzovaly dopad parazita na jejich výkonnost.
2.3.1.3. Adaptace parazita
Parazité jsou velmi dobře adaptováni k přenosu mezi jednotlivými hostiteli. Mezi takové adaptace patří např. zvyšování počtu potomků nebo odolnost přenosných stádií proti působení vnějšího prostředí. Parazit také může podpořit přenos do dalšího hostitele indukcí změn v chování hostitele zvyšujících pravděpodobnost takového přenosu. Podle Dawkinse (1998) může být fenomén vyskytující se mimo tělo organismu rozšířeným fenotypickým projevem genotypu organismu. Je možné potom hovořit o některých změnách v chování hostitele jako o adaptaci parazita. Ukazuje se, že množství změn v chování parazitovaných jedinců skutečně zvyšuje pravděpodobnost přenosu parazita do dalšího hostitele. Je však velmi obtížné rozhodnout, zda-li pozorované změny v chování jsou skutečně produktem specifické manipulace hostitele parazitem nebo vedlejším efektem onemocnění. Poulin (1995) vymezuje několik kritérií, které by měly změny v chování splňovat, abychom mohli hovořit o těchto změnách jako o adaptaci parazita:
Změny v chování hostitele by měly být z hlediska parazita účelné. Například u parazitů přenosných predací, by měly pozorované změny zvyšovat pravděpodobnost ulovení mezihostitele predátorem - konečným hostitelem.
Dochází-li u hostitele následkem parazitace ke komplexním změnám v chování či vzniku zcela nových typů chování, je nepravděpodobné, že vznikly jako důsledek náhody. Komplexní změny v chování hostitele naznačují, že by se mohlo jednat o specifické působení parazita spíše než o vedlejší efekt onemocnění.
Vyvolávají-li nepříbuzní parazité se shodným způsobem přenosu podobné změny v chování čili dochází ke konvergenci, je to dalším argumentem podporujícím přítomnost manipulační aktivity.
Důležitým kritériem k posouzení toho, že dané chování je z hlediska parazita adaptivní, je zvyšování fitnes parazita. Měřit fitnes je však velmi obtížné. U parazitů přenosných predací jsou jediným skutečným ověřením adaptivnosti pozorovaných změn v chování hostitele, predační pokusy (Poulin 1995).
2.3.1.3.1. Možnosti manipulace hostitele parazitem
Účelem manipulace hostitele je zvýšení efektivnosti přenosu parazita. Případná manipulace bude zaměřena na takový typ chování, který bude zvyšovat pravděpodobnost transmise. Podle způsobu přenosu můžeme tedy očekávat změny v určitém typu chování.
Sexuálně přenosní parazité by mohli ovlivňovat sexuální chování hostitele. Protože vyšší sexuální aktivita může být i v zájmu hostitele, čili nedochází v tomto případě ke konfliktu zájmů, je velmi obtížné rozhodnout zda se jedná o příčinu nebo důsledek infekce. Změny v sexuálním chování způsobené pohlavně přenosným parazitem nebyly doposud zjištěny.
Parazit může měnit disperzi hostitele. Pokud se nepřekrývá areál výskytu hostitele v němž se parazit nachází s arálem výskytu následného hostitele, může parazit manipulovat hostitele ke vstupu do lokalit, kde se vyskytuje následný hostitel. Např. vodní měkkýši Illianasa adunca infikovaní motolicí Gynaecotyla adunca se obvykle zdržují v blízkosti pobřeží, na rozdíl od neparazitovaných jedinců žijících ve větší hloubce. Parazitovaní měkkýši se tak dostávají do míst, v nichž se vyskytuje další mezihostitel motolice - semiterestriální korýš (Curtis 1990 dle Moore 1995).
Nejvíce změn v chování hostitele o nichž se uvažuje jako o adaptaci parazita, bylo popsáno u parazitů přenosných predací. Holmes a Bethel (1972) naznačují čtyři způsoby jimiž může parazit měnit vztah mezi kořistí a predátorem a zvyšovat tím pravděpodobnost přenosu:
Přestože autoři studií se velmi často domnívají, že změny v chování hostitele zvyšují náchylnost k predaci, k skutečnému ověření jsou nutné predační testy (Poulin 1995).
2.3.2. Změny v aktivitě
Behavaiorální efekt parazitace má u zvířat velmi často podobu změn v aktivitě nakaženého jedince. Podobně i u člověka je infekce některými parazity spojena se změnami ve fyzické aktivitě.
Kvalsvig (1998) sledovala vliv schistomiasy (Schistosoma haematobium, Schistosoma mansoni) na aktivitu při volné hře u dětí z jihoafrického venkova. Lehká infekce (měřeno počtem vajíček) neměla vliv na aktivitu dětí. U dětí s vyšší intenzitou infekce nebo s kombinovanou infekcí oběma druhy schistosom byla zjištěna nižší aktivita při hře projevující se obzvláště za horkého počasí. Intervenční studií (Kvalsvig 1981) bylo zjištěno, že léčba schistosomiasy vede ke zvýšení aktivity dětí ve srovnání s neléčenými kontrolami. Typickým důsledkem schistosomiasy je letargie, která však může být maskována selektivní exposicí aktivnějších dětí k onemocnění (Kvalsvig 1998).
Léčba dětí s jednoduchou nebo kombinovanou infekcí helminty Ascaris lumbricoides, Trichuris trichiura a Ancylostoma duodenale vedla k zvýšení fyzické aktivity při volné hře v porovnání s placebo skupinou. Největší efekt na zvýšení aktivity mělo snížení intenzity infekce Trichuris trichiura (Adams a spol. 1994).
Connolly a Kvalsvig (1992) se domnívají, že společným důsledkem parazitárních infekcí je letargie. Hostitel je netečný, pomalý v akci a celkově se mu nedostává energie, což se projeví nejen ve fyzické aktivitě, ale i v mentální činnosti.
U zvířat bylo zjištěno, že někteří parazité (např. Toxocara canis u myši) způsobují hyperaktivitu hostitele. Přestože se v případě toxokariasy uvažuje o stejném efektu i u člověka (Hay a spol. 1986), nebyly doposud provedeny žádné studie, které by u člověka zjistily kauzální souvislost mezi parazitací a hyperaktivitou.
2.3.3. Změny kognitivních funkcí
Dá se předpokládat, že infekční onemocnění bude mít dopad nejen na fyzickou, ale i mentální výkonnost jedince. Nicméně poměrně málo studií se zabývalo dopadem jednotlivých infekčních agens na konkrétní kognitivní procesy.
Efekt experimentálně navozené infekce bakterií Pasteurella tularensis sledovali Alluisi a spol. (1971). Tento patogen způsobuje tularemii, onemocnění charakterizované těžkými bolestmi hlavy, fotofobií, myalgií, nevolností a depresí. U infikovaných osob došlo k průměrnému zhoršení výkonnosti v baterii testů o 25% oproti preinfekční výkonnosti. Jednotlivé testy byly určeny k hodnocení pozornosti, paměti, senzorické percepce a procedurálních operací. Bylo však hodnoceno pouze celkové skóre, takže není možné rozhodnout, které funkce byly infekcí ovlivněny.
Behaviorální změny byly studovány také u některých virových infekcí. Onemocnění infekční mononukleosou má vliv jak na náladu tak i na výkonnost. Infikované osoby vykazovaly vyšší míru úzkosti a deprese. Osoby v akutní fázi infekce měly pomalejší reakční rychlost a hůře udržovaly pozornost ve srovnání s kontrolami. U osob v chronické fázi infekce bylo zjištěno zhoršení krátkodobé paměti a logického zdůvodňování. Zatímco afektivní změny korelovaly s intenzitou symptomů, signifikantní vztah mezi výsledky výkonnostních testů a intenzitou symptomů zjištěn nebyl (Hall & Smith 1996a).
Podobný selektivní efekt na výkon jako u infekční mononukleosy byl zjištěn také u experimentální nákazy viry způsobujícími onemocnění horních cest dýchacích (rýmu) a chřipku. Onemocnění horních cest dýchacích mělo vliv na reakční čas a tento efekt byl pozorován i po zmizení klinických symptomů. Oproti tomu onemocnění chřipkou zhoršovalo výkon v úkolech, při nichž testovaný subjekt neví, kde se cílový stimulus objeví (Smith a spol. 1989). Vzhledem k tomu, že tato práce byla založena na velmi malém počtu osob (5 a 10) je otázkou jak jsou její výsledky průkazné (Savory 1992). Na druhou stranu podobný efekt byl zjištěn i u přirozeně se vyskytujícího onemocnění horních cest dýchacích. Kromě zhoršení reakčního času, bylo zjištěno i zhoršení v úkolech vyžadujících udržení pozornosti (pouze v druhém týdnu) a vzrůst negativní nálady v prvním týdnu po nákaze (Hall & Smith 1996b). Mechanismus odpovědný za pozorované behaviorální změny není znám. Hall a Smith (1996a) se domnívají, že produkce cytokinů jako součást imunitní reakce na virovou infekci má přímý efekt na centrální nervovou soustavu.
Také infekce některými helminty je spojena se změnami v kognitivních funkcích. Děti se střední mírou infekce tenkohlavcem Trichuris trichiura dosahovaly horších výsledků v testech z aritmetiky a čtení oproti nenakaženým dětem. Proměnné, které ovliňovaly výkon jako socioekonomický status, školní docházka, věk a pohlaví, byly kontrolovány (Simeon a spol. 1994). Léčba vedla u dětí s těžkou mírou infekce k zlepšení v gramatických testech oproti dětem dostávajícím placebo. V testech z aritmetiky a čtení nebyl efekt léčby zjištěn. U dětí s lehkou formou nákazy se po léčbě výkon v jednotlivých testech, ve srovnání s neléčenými dětmi, výrazně nezlepšil (Simeon a spol. 1995). Nokes a spol. (1992) zjistili, že odstranění parazita anthelmiky vedlo po devíti týdnech k signifikantnímu zlepšení krátkodobé sluchové paměti a vybavování dlouhodobé paměti. Nebyla však nalezena korelace mezi intenzitou infekce a zlepšením v testech.
Jiným značně rozšířeným helmintem, jenž má vliv na kognitivní funkce hostitele, je škrkavka Ascaris lumbricoides. Léčba askariazy vedla po pěti měsících ke zlepšení v testu měřícím motorickou koordinaci, rychlost mentálních operací, krátodobou paměť a v Ravenově barevném testu (měří schopnost zdůvodňovat) (Hadidjaja a spol. 1998). Intenzita infekce je negativně korelována s verbálními schopnostmi a inhibičně kontrolním aspektem kognitivního chování. Mezi nakaženými dětmi se také častěji vyskytovaly abnormality v EEG (Levav a spol. 1995). Většina infikovaných dětí (86%) však byla v případě této studie postižena malnutricí. Nutriční status dětí měl také rozhodující vliv na výkon v kognitivních testech.
Jaké konkrétní kognitivní funkce jsou helmintárními infekcemi ovlivněny, není možné podle doposud provedených studií rozhodnout. Jednak bylo v některých studiích použito k hodnocení kognitvních funkcí školních znalostí, které je kvůli jejich komplexnosti velmi obtížné interpretovat. Školní výsledky jsou ovlivněny množstvím faktorů jako je např. dostupnost přiměřené výuky, kvalita výuky, dostupnost školních pomůcek, socioekonomický staus rodičů, množství absencí aj. Některé z nich byly v uváděných studiích (např. školní docházka) kontrolovány. Je však v podstatě nereálné kontrolovat všechny faktory ovlivňující školní výkon, a je proto vhodnější používat testy konkrétních kognitivních funkcí. Dalším problémem s interpretací výsledků je, že v jednotlivých studiích bylo použito různých testů, což omezuje jejich vzájemné porovnání.
Není také znám mechanismus, jímž uváděné parazitární infekce ovlivňují kognitivní funkce člověka. K ovlivnění by mohlo docházet několika různými způsoby. Parazit může způsobovat přímé biochemické změny v centrálním nervovém systému. Například infekce Trichiuris trichiura způsobuje deficienci železa (Solomons 1993). Nedostatek železa vede k behaviorálním změnám v důsledku změn metabolismu katecholaminů (Connolly & Kvalsvig 1993). K ovlivnění kognitivních funkcí by mohlo docházet i nepřímo, např. změnou nutričního stavu jedince. Onemocnění askariasou i trichiurasou má za následek poruchy zažívání a vede i k malnutrici (Solomons 1993). Je známo, že jedním z důsledků podvýživy je i zhoršení kognitivních funkcí (Connolly & Kvalsvig 1993). Nutriční stav může tedy vystupovat, jednak jako rušivá proměnná, ale taktéž jako mechanismus, jímž infekce může ovlivnit mentální výkon. Na druhou stranu dopad infekce na kognitivní funkce byl zjištěn i v souborech, kde se malnutrice a anémie vyskytovaly jen zřídka (Simeon a spol. 1994; Nokes a spol. 1992).
Horší kognitivní výkony u nakažených dětí mohou být také způsobeny snížením aktivity a tím redukcí stimulace a možností učení (Nokes & Bundy1994).
2.3.4. Behaviorální efekt toxoplasmosy
2.3.4.1. Změny v chování hlodavců
Přenos parazita do konečného hostitele - kočky probíhá, pomineme-li možnou nákazu oocystami od jiné kočky, obvykle formou predace. Přirozenou kořistí kočky jsou drobní hlodavci. Zvýšení pravděpodobnosti přenosu do kočky by mohl parazit dosáhnout indukcí chování zvyšující pravděpodobnost ulovení hlodavce kočkou.
Behaviorální efekt toxoplasmosy byl nejčastěji studován u laboratorní myši. Hay a spol. (1983a) zjistili zhoršené motorické schopnosti, měřené počtem pádů z rotujícího válce, u nakažených myší oproti kontrolám. Myši nakažené kongenitálně padaly signifikantně častěji než myši nakažené v dospělosti a obě skupiny byly signifikantně horší než skupina kontrolní. Zhoršená motorická výkonnost nebyla ovlivněna počtem tkáňových cyst v mozku ani zdravotním stavem nakažených zvířat.
Neofilie patří u myší mezi typické prvky chování. Pokud se myš dostane do nového prostředí stráví množství času jeho prozkoumáváním. Pozorováním infikovaných myší v novém prostředí byl zjištěn nižší výskyt exploračního chování jako je např. panáčkování (Hutchison a spol. 1980b). Mimo to se u nakažených zvířat objevovalo častější střídání jednotlivých prvků chování. Nakažené myši vykazovaly relativně nižší preferenci pro centrální oblast boxu (Hay a spol. 1983b,1984). Při pokusu v Y bludišti dávaly kontrolní myši přednost novému ramenu zatímco u nakažených myší nebyla zjištěna žádná preference (Hutchison a spol. 1980a). Vypadá to tedy, že infekce Toxoplasma gondii ovlivňuje u myší schopnost rozlišovat mezi novými a známými podněty (Hutchison a spol. 1980b). Několik prací se věnovalo změnám v aktivitě nakažených myší. Myši infikované kongenitálně i v dospělosti byly oproti kontrolám významně aktivnější, a to jak v novém prostředí (Hay a spol. 1983b), tak i v prostředí domácí klece (testovány byly pouze kongenitálně nakažené myši) (Hay a spol. 1985). Ve všech případech nebyla změna v aktivitě korelována s počtem tkáňových cyst v mozku ani celkovým zdravotním stavem. Podle Hay a spol. (1983a) jsou změny v chování infikovaných myší důsledkem histopatologických změn v mozku způsobených tkáňovými cystami. Zitková (1996) sledovala dynamiku změn v aktivitě nakažených myší v průběhu dvanácti týdnů od nákazy. Zjistila nižší aktivitu u nakažených zvířat v akutní fázi infekce, při níž byly u všech nakažených pozorovány známky onemocnění. V chronické fázi infekce (testováno 6. a 12. týden po infekci) se aktivita nakažených myší od kontrol již nelišila. Autorka se na základě svých výsledků domnívá, že změny v aktivitě jsou pouze přechodné a jde o vedlejší produkt onemocnění a nikoliv o manipulační aktivitou parazita. V oblasti sociálních interakcí byli kongenitálně nakažení samci více agresivní a častěji zkoumali jiné jedince (Arnott a spol. 1990). Zitková (1996) však zvýšení agresivity u nakažených samců nepozorovala.
Změny v chování byly pozorovány také u potkana Rattus norvegicus. Pro chování potkanů je na rozdíl od myší typická neofobie. Nakažení potkani vykazovali nižší míru neofobie. Oproti kontrolám dříve ztráceli obavu žrát z misky s novým podnětem (pach, miska, potrava). Pokud byla potrava pouze v pastech, chytali se do nich nakažení jedinci dříve než jedinci kontrolní (Webster a spol. 1994). Infekce Toxoplasma gondii měla také vliv na vyšší aktivitu nakažených zvířat (Webster 1994). Výsledky těchto prací jsou o to závažnější, že stejný efekt vykazovaly skupiny s experimentálně nakaženými hybridy divokého a laboratoního potkana tak i odchycení divocí potkani nakažení přirozenou formou. V oblasti sociálního chování pozorovaného v přirozených podmínkách výběhu neměla nákaza vliv na sociální status ani na kopulační chování. Infikovaní jedinci však oproti kontrolám signifikantně častěji explorovali nové podněty (Berdoy a spol. 1995). Podle autorů všechny pozorované změny v chování nakažených potkanů zvyšují pravděpodobnost, že budou uloveni kočkou. Protože se jedná o změny specifické, domnívají se, že jde spíše o manipulaci hostitele ze strany parazita než o vedlejší produkt onemocnění.
Výsledky behaviorálních pokusů podporuje i zjištění změn v koncetracích neurotransmiterů u infikovaných myší oproti kontrolám. U myší s akutní toxoplasmosou byl zjištěn vzrůst kyseliny homovanilikové o 40% a pokles norepinefrinu o 28%, hladina dopaminu, serotoninu a kyseliny 5-hydroxyindolacetové zůstala nezměněna. Ve skupině myší s chronickou toxoplasmosou byla hladina dopaminu o 14% vyšší, hladiny ostatních neurotransmiterů zůstaly nezměněny (Stibbs 1985). Tyto látky zprostředkují lokomoční aktivitu, náladu, agresi, spánek, učení, paměť a průtok krve mozkem. Tyto neurochemické změny mohou být faktorem přispívajícím k mentálním a motorickým abnormalitám spojených nebo způsobených toxoplasmosou u hlodavců a možná i u člověka (Stibbs 1985).
Pro rozhodnutí, zda u pozorovaných změn v chování parazitovaných jedinců jde skutečně o adaptaci parazita, jsou nezbytné predační pokusy (Poulin 1995), ty však v případě jedinců nakažených Toxoplasma gondii nebyly doposud provedeny.
2.3.4.2. Vliv toxoplasmosy na vznik psychiatrických poruch
Časté poruchy chování u pacientů s cerebrální formou toxoplasmosy vedly k diskusím o podílu toxoplasmosy na vzniku psychických onemocnění. Jírovec a Vojtěchovský (1955) zjistili u pacientů z psychiatrických léčeben positivní reakci na toxoplasminový intradermální test v 54,5%, 50% a 40% případů, oproti 30% pozitivních reakcí v normální populaci. Podobné výsledky, 59,3% pozitivně reagujících psychiatrických pacientů, získali i Petrovický a Kvapilík (1955). U obou studií tvořili více jak polovinu pacientů schizofrenici. U některých pacientů s těžkou formou akutní toxoplasmosy byl zaznamenán vznik schizofrenního procesu (Kretschmer & Schmid 1955; Minto & Roberts 1959). Petrovický a Vojtěchovský (1955) se domnívají, že toxoplasmosa narušila, nebo u některých nemocných stále narušuje centrální nervový systém a činí jej tak více náchylný k vzniku duševního onemocnění. Ladee (1966) však upozorňuje, že je obtížné rozlišit, zda je toxoplasmosa faktorem vyvolávajícím nebo faktorem pouze přídavným při vzniku schizofrenního procesu. Autor popisuje dva případy schizofrenie u nichž, přestože byly zjištěny vysoké titry protilátek proti Toxoplasma gondii, nebyla toxoplasmosa etiologicky korelována se schizofrenním procesem. Nejčastější forma toxoplasmosy je však asymptomatická a těžké formy akutní toxoplasmosy, jejichž následkem mohou být i psychiatrické poruchy, se u imunokompetentních osob vyskytují vzácně. Vysoká prevalence u pacientů v psychiatrických léčebnách je tedy spíše faktorem pouze koincidujícím a celkový podíl toxoplasmosy na vznik schizofrenních onemocnění je nevýznamný.
Hoschl a Balon (1980) porovnali dotazníkovou metodou skupinu pacientů léčených na toxoplasmosu a skupinu pacientů psychiatrické léčebny z oddělení neuroz. Skupina pacientů z oddělení neuroz vykazovala významně vyšší míru neurotičnosti a životní frustrace. Přesto bylo i u pacientů s toxoplasmosou zjištěno množství neurotických symptomů a lze je považovat za součást klinického obrazu toxoplasmosy. Vzhledem k tomu, že serologický test na toxoplasmosu byl ve skupině neurotiků ve všech případech negativní, je zřejmě podíl toxoplasmosy na vznik neurotického onemocnění nevýznamný.
Také v ústavech pro mentálně postižené děti byla zjištěna vyšší prevalence toxoplasmosy oproti normální populaci (Robertson 1965). Tyto nálezy byly dávány do souvislosti s kongenitální toxoplasmosou (Thalhammer 1962). Jiné studie však zjistily prevalenci toxoplasmosy u mentálně postižených stejnou (Mackie a spol. 1971) nebo dokonce i nižší než v normální populaci (Cook & Derrick 1961). Burkinshaw a spol. (1953) vyšetřovali na toxoplasmosu mentálně postižené v Anglii a zjistili, že v žádném z vyšetřovaných případů nelze mentální deficienci definitivně přičítat toxoplasmose. Podle Garcia (1979) je vyšší prevalence mezi mentálně postiženými způsobeno jejich horšími hygienickými návyky.
2.3.4.3. Změny osobnosti
Následkem infekce parazitem Toxoplasma gondii může docházet také ke změnám v osobnosti člověka. Jak již bylo uvedeno, může docházet u některých pacientů s cerebrální formou akutní toxoplasmosy k patologickým změnám osobnosti (Freytag & Haas 1979). Zda-li má vliv na osobnost také chronická forma toxoplasmosy, která je ve většině případů asymptomatická, studovali Flegr a Hrdý (1994) a Flegr a spol. (1996). V případě první studie se experimentální soubor skládal z 328 (195 mužů a 143 žen) dobrovolníků z řad univerzitních studentů a pedagogů biologie. Existence specifické buněčné imunity byla zjištěna na základě intradermálního testu. Na zjišťování osobního profilu byl použit Cattellův šestnáctifaktorový dotazník. Mezi skupinou Toxoplasma pozitivních a Toxoplasma negativních mužů byly zjištěny signifikantní rozdíly ve faktorech G (síla superega) a L (protense). Mezi nakaženými a nenakaženými ženami nebyly signifikantní rozdíly v osobnostním profilu zjištěny. V následující studii byl sledovaný soubor rozšířen na 394 osob (224 mužů a 170 žen). Mezi skupinou nakažených osob byly opět zjištěny rozdíly v některých faktorech osobnosti monitorovaných Cattellovým dotazníkem, a to jak u mužů, tak i u žen. Bylo zjištěno, že u mužů je infekce spojena, stejně jako v první studii, s nízkou sílou superega (nespolehlivý, lehkovážný) a protensí (podezíravý, žárlivý) a dále s vyšším skórem ve faktoru O (náchylnost k pocitům viny) a nižším skórem ve faktoru Q2 (závislost na skupině). U žen je infekce Toxoplasma gondii charakterizována vysokým skórem ve faktorech A (afektothymie - vřelý, společenský) a Q2 (soběstačnost) a nízkým skórem ve faktorech L (důvěřivý, přizpůsobivý) a O (klidná sebedůvěra). Ve faktorech L, O, Q2 má tedy infekce opačný efekt u mužů než u žen.
Rozdíly v osobnostním profilu byly také zjištěny v náhodném vzorku 191 žen z normální české populace (Flegr & Havlíček 1999). Tyto rozdíly však nebylo možné pozorovat přímo. Ženy byly testovány na toxoplasmosu během těhotenství a osobnostní dotazník jim byl zasílán poštou, nebylo jim však známo, že se studie týká toxoplasmosy. Na základě běžného serologického vyšetření (komplement fixační reakce) bylo 136 žen hodnoceno jako Toxoplasma negativní a 55 žen Toxoplasma pozitivní. Mezi skupinami vytvořenými podle výsledků serologického testu nebyly zjištěny žádné rozdíly v osobnostním profilu. Tato serologická metoda je však vhodná především k hodnocení akutní toxoplasmosy, na rozdíl od intradermálního testu použitého u souboru biologů, a některé osoby nakažené před dlouhou dobou mohou být hodnoceny jako falešně negativní. Protože případné změny v osobnosti by se projevily až určitou dobu po nákaze, je možné, že osoby s velmi starými infekcemi, a tedy s největším rozsahem změn, byly serologickým testem hodnoceny jako falešně negativní a tím byly zastřeny i rozdíly mezi oběma skupinami. Na eliminaci efektu kontaminace dat byl použit permutační test. Tímto testem bylo zjištěno, že nakažené ženy v porovnání s nenakaženými ženami dosahovaly vyššího skóre ve faktorech B (inteligence), O (náchylnost k pocitům viny) a dále také ve faktorech Q3 (radikalismus) a Q4 (vysoká ergická tenze). Poněkud rozdílné výsledky získané v souboru biologů jsou zřejmě důsledkem rozdílnosti částí populace, z nichž se tyto soubory rekrutovaly.
Zjištěné asociace mezi infekcí toxoplasmou a osobnostním profilem mohou být buď důsledkem parazitace a nebo určité faktory osobnosti zvyšují pravděpodobnost infekce parazitem. Pokud by docházelo k indukci změn parazitem, projeví se tyto změny až určitou dobu po nakažení, čili měla by existovat korelace mezi délkou infekce a intenzitou pozorovaných změn (Flegr a spol. 1996). Za účelem rozhodnutí mezi těmito dvěma mechanismy byl zasílán Cattellův dotazník mužům, u nichž byla během minulých třinácti let diagnostikována akutní toxoplasmosa. Na základě 164 získaných osobnostních dotazníků a klinických záznamů bylo z šestnácti monitorovaných faktorů zjištěno systematické snižování skóre ve faktoru G (nízká síla superega) v závislosti na délce infekce. Jedná se o stejný faktor, jehož nízká hladina byla zjištěna ve skupině Toxoplasma pozitivních biologů (Flegr a spol. 1996).
Korelace mezi délkou infekce a systematickými změnami byla také zjištěna u dvou nezávislých souborů žen. Ve skupině 230 žen, u nichž byla známa délka infekce, byl zjištěn systematický vzestup ve faktorech G (síla super ega) a Q3 (sebekontrola). V druhém souboru 55 žen byla délka infekce odhadována podle hladin titrů protilátek (u starších infekcí jsou nižší). Opět byla zjištěna korelace mezi délkou infekce a vzestupem ve faktoru G a A (afekthotymie) (Flegr a spol. zasláno do tisku). S délkou infekce tedy korelovaly stejné faktory, v kterých byl zjištěn rozdíl mezi nakaženými a nenakaženými ženami ze souboru biologů.
Tyto výsledky tedy naznačují, že změny osobnosti jsou spíše indukovány parazitem, než že určitá kombinace faktorů osobnosti zvyšuje pravděpodobnost infekce. Jestli skutečně dochází k systematickým změnám v osobnosti, je však třeba ověřit formou test-retest v rozmezí více let, aby bylo možné případné změny identifikovat.
3. Cíle práce
Tato práce se zabývá dopadem latentní toxoplasmosy na lidský organismus. Část práce navazuje na předešlé studie (Flegr & Hrdý 1994; Flegr a spol. 1996) v nichž byly zjištěny rozdíly v profilu osobnosti mezi skupinou nakažených a nenakažených osob. Zjištěné rozdíly mohou být důsledkem dvou rozdílných mechanismů. Jedná se buď o důsledek působení parazita, anebo určité faktory osobnosti zvyšují riziko infekce. Pokud existují asociace mezi epidemiologickými faktory a faktory osobnosti, v nichž byly zjištěny rozdíly mezi nakaženými a nenakaženými osobami, jsou tyto rozdíly daleko spíše příčinou infekce. Naproti tomu pokud by zjištěné rozdíly v osobnostních faktorech byly výsledkem působení parazita, mělo by u těchto faktorů docházet k systematickým změnám. Na rozhodnutí mezi těmito dvěma mechanismy bylo zaměřeno několik dílčích cílů práce. Prvním úkolem bylo zjistit, které epidemiologické faktory zvyšují riziko infekce Toxoplasma gondii. Dalším úkolem bylo zjistit zda-li existují asociace mezi faktory osobnosti a epidemiologickými faktory u nichž bylo zjištěno, že zvyšují riziko infekce. Existence systematických změn v osobnosti způsobených parazitem byla zjišťována retestem po pěti letech od původního testování. Posledním cílem této části práce bylo ověřit existenci rozdílů v osobnostním profilu na nezávislém souboru.
Další část práce byla zaměřena na výkon v psychomotorických testech. U některých virových onemocnění např. u infekční mononukleózy je známo, že způsobují zhoršení výkonnosti v psychomotorických testech a to i poté co odezní klinické příznaky onemocnění (Hall & Smith 1996a). Efekt latentní toxoplasmosy na psychomotorický výkon není znám. Cílem této části práce bylo zjistit vliv latentní toxoplasmosy na psychomotorickou výkonnost v testech reakční rychlosti. Pokud by byly zaznamenány změny v reakčním čase v důsledku infekce, bylo by zajímavé vědět, zda-li se tyto změny uplatňují také v situacích běžného života. Takovou situací v níž se reakční čas uplatňuje jsou i dopravní nehody. Cílem této části práce bylo zjistit jaká je prevalence toxoplasmosy u osob, které se staly účastníky dopravní nehody jako chodci nebo jako řidiči.
Poslední část práce byla zaměřena na dopad latentní toxoplasmosy na lidské zdraví. Přestože tento parazit patří u člověka mezi nejrozšířenější infekční agens, následkům dlouhodobé infekce bylo věnováno poměrně málo pozornosti. Latentní toxoplasmosa je obvykle považována za asymptomatickou, s minimem klinických následků. Cílem této části práce bylo zjistit zda má latentní toxoplasmosa vliv na subjektivní hodnocení zdravotního stavu. V poslední době bylo zjištěno, že latentní toxoplasmosa je spojena s nižší tělesnou hmotností (Flegr & Hrdá v tisku). Hmotnost je však nutné vždy hodnotit ve vztahu k výšce např. formou Body Mass Indexu (BMI). Cílem této části práce bylo zjistit vliv latentní toxoplasmosy na index tělesné hmotnosti (BMI).
3. Materiál a metody
3.1. Soubory osob
V celé práci byly použity tři nezávislé soubory osob, pro něž jsou dále použita tato označení: biologové, dárci krve a pacienti.
Soubor biologů
Soubor se skládal ze 414 osob (233 mužů a 186 žen), studentů a pedagogů z biologické sekce na Př.F.U.K. Osoby byly v rámci předešlých studií (Flegr & Hrdý 1994; Flegr a spol. 1996) vyšetřeny na toxoplasmosu intradermálním testem a většina z nich vyplnila i Cattellův šestnáctifaktorový osobnostní dotazník. Celkem 379 osobám u nichž byla známa adresa, byl poštou zaslán epidemiologický dotazník. Psychomotorické testy podstoupilo 117 (66 mužů a 52 žen) osob. Osoby byly vybrány podle náhodné dosažitelnosti v období provádění testů v budově univerzity či na kolejích. Osobnostní dotazník byl zaslán po pěti letech od původního vyplňování celkem 373 osobám.
Soubor dárců krve
Tento soubor se skládal ze 127 dobrovolných dárců trombocytů na Ústavu hematologie a krevní transfúze při Fakultní nemocnici 1. Lékařské fakulty Univerzity Karlovy. V průběhu odběru trombocytů na destičkovém separátoru byli dárci tázáni, zda-li jsou ochotni podstoupit test na reakční rychlost. Kromě testu reakční rychlosti byla také změřena jejich tělesná výška a hmotnost. Ke konci vyšetření byly osoby ještě požádány, zda-li by nemohly doma vyplnit Cattellův osobnostní dotazník a v přiložené obálce ho zaslat zpět. V době vyšetřování a vyplňování dotazníku nebyly probandům ani administrátorovi známy výsledky seorlogických testů na toxoplasmosu.
Soubor pacientů
Soubor byl složen ze 169 pacientů z chirurgického oddělení Všeobecné fakultní nemocnice Na Královských Vinohradech. Osoby byly přivezeny na oddělení poté, co se staly účastníky dopravní nehody. Podle lékařského záznamu bylo zjištěno jakým způsobem se pacient na dopravní nehodě podílel. Serologické testy na toxoplasmosu byly provedeny v Národní referenční laboratoři pro toxoplasmosu při Státním zdravotním ústavu.
3.2. Epidemiologický dotazník
Dotazník byl osobám zasílán poštou s žádostí o co nejrychlejší odpověď. Byly v něm položeny otázky týkající se faktorů, které mohou zvýšit riziko infekce toxoplasmou (konzumace syrového masa, kontakt s kočkou, chov psa a králíků) a dále otázky ohledně zdravotního stavu. Z hlediska získání toxoplasmosy je jedním z rizikových faktorů i život na venkově. Vzhledem k tomu, že incidence toxoplasmosy je u nás nejvyšší ve věkových kategoriích do dvaceti let, je důležité znát, kde studovaná osoba prožila toto období. Tato otázka byla položena v rámci Cattellova osobnostního dotazníku rozesílaného probandům po pěti letech od původního vyplňování. Na jednotlivé otázky bylo možno odpovědět buď pouze „ano-ne“ nebo byly k otázce připojeny možné odpovědi. Přesné znění epidemiologických otázek a nabízených odpovědí je uvedeno v tab. č.2.
Máte (měli jste) doma (na chalupě) kočku?
Jak často hladíte kočku (nikdy, jednou za pět let, za dva roky, za rok, za měsíc, častěji)?
Máte (měli jste) doma (na chalupě) psa?
Máte (měli jste) doma (na chalupě) králíky?
Jedl(a) jste (nebo ochutnával při vaření) syrové maso?
Mládí jste prožil(a) převážně (na venkově, ve městě nad pět tisíc obyvatel, ve městě nad sto tisíc obyvatel).
Tab. č. 2: Přehled epidemiologických otázek a nabízených odpovědí zasílaných probandům poštou. Poslední otázka byla položena v rámci Cattellova osobnostního dotazníku.
3.3. Dotazník na subjektivní hodnocení zdravotního stavu
Subjektivní hodnocení zdravotního stavu bylo zjišťováno dotazníkovou metodou. Osoby byly tázány na hodnocení svého fyzického zdraví, psychické pohody, zda trpí alergickými chorobami a na frekvenci běžných onemocnění (chřipka, angína). Otázky byly položeny v rámci epidemiologického dotazníku rozesílaného poštou. Možné odpovědi byly pouze „ano-ne“ a u otázky na frekvenci onemocnění byly nabídnuty možné odpovědi. Přesné znění všech otázek a nabízených odpovědí je uvedeno v tab. č. xx.
Jak často onemocníte běžnými chorobami (chřipka, angína) (vůbec ne, jednou za pět let, za dva roky, za rok, častěji)?
Trpíte alergickými chorobami (např. astma, senná rýma)?
Cítíte se obvykle dobře a v duševní pohodě?
Cítíte se obvykle fyzicky zdráv?
Tab. č. xx: Přehled otázek a nabízených odpovědí týkajících se zdravotního stavu položených v rámci epidemiologického dotazníku
3.4. Cattellův osobnostní dotazník
K zjišťování osobnostního profilu byl použit Cattellův šestnáctifaktorový dotazník, forma A. Tento dotazník patří k často využívaným metodám v psychologické diagnostice a výzkumu v České republice i v zahraničí (Říčan 1975). Profil osobnosti je charakterizován šestnácti faktory (viz. tab. č. xx) na základě odpovědí na 187 položek (otázek). Výsledkem dotazníku je pro každý faktor hrubé skóre na dvacetijedna stupňové případně dvacetisedmi stupňové škále. Hrubá skóre jsou v psychologické diagnostice obvykle transformována do desti stupňové věkově standartizované škály. Protože soubory osob jimž byly dotazníky administrovány (biologové, dárci krve) jsou představitelé spíše netypické subpopulace, byla kvůli zamezení vlivu věkových korekcí určených pro obecnou populaci, k hodnocení použita původní hrubá skóre. Efekt věku byl odfiltrován jiným způsobem (viz. kap. 4.8.). Biologům, kteří vyplnili dotazník v rámci předešlých studií (Flegr & Hrdý 1994; Flegr a spol. 1996), byl po pěti letech od původního testování, tzn. v letech 1997-99, zaslán dotazník poštou. V průvodním dopise byly osoby informovány, že se jedná o pokračování studie jíž se před pěti lety zúčastnily a byly požádány o další spolupráci. Dárcům krve byl dotazník předán při psychomotorickém testování s žádostí o vyplnění a zaslání zpět přiloženou obálkou. Osoby byly pouze informovány, že se jedná o součást diplomové práce, účel studie jim však nebyl znám.
|
faktor |
nízké skóre |
vysoké skóre |
|
A |
Sizotýmie rezervovaný, kritický, uzavřený |
Afektotýmie vřelý, přátelský, bezstarostný, společenský |
|
B |
Nízká inteligence |
Vysoká inteligence |
|
C |
Slabost ega ovlivňován city, emočně méně stálý |
Vyšší síla ega emočně stálý, zralý, čelí realitě, klidný |
|
E |
Submisivnost poslušný, mírný, přizpůsobuje se |
Dominance prosazuje se, je agresivní, soutěživý, |
|
F |
Desurgence střízlivý, mlčenlivý, vážný |
Surgence entuziastický, bezstarostný |
|
G |
Malá síla superega nedodržuje pravidla,nesvědomitý, nestálý |
Síla superega svědomitý, vytrvalý, moralista, spolehlivý |
|
H |
Threctia plachý, ostýchavý, citlivý k hrozbě |
Parmia dobrudružný, společensky smělý |
|
I |
Harria houževnatý, tvrdý, realista |
Premsia sensitivní, jemný, závislý, rozmazlený |
|
L |
Alaxia důvěřivý, přijímá dané podmínky |
Protense podezíravý, žárlivý |
|
M |
Praxernia praktický, má „přízemní“ zájmy a starosti |
Autia imaginativní, bohémský |
|
N |
Naivita upřímný, neokázalý |
Sofistikovanost protřelý, „znalý světa“ |
|
O |
Klidná sebedůvěra sebejistota, radostnost, spokojenost |
Sklon k pocitům viny pln obav a starostí, pohrdá sebou, nejistý |
|
Q1 |
Konservativní povaha konzervativní, respektuje uznávané myšlenky |
Radikalismus experimentující, liberální, volné myšlení |
|
Q2 |
Závislost na skupině sociabilně závislý na skupině |
Soběstačnost soběstačný, rád se samostatně rozhoduje |
|
Q3 |
Nízká integrace selfsentimentu bez sebekontroly, laxní, řídí se svými popudy |
Velká síla selfsentimentu sebekontrola, síla vůle, řídí se svým sebepojetím |
|
Q4 |
Nízká ergická tenze relaxovaný, poklidný, uspokojený |
Vysoká ergická tenze napjatý, frustrovaný, podrážděný, popudlivý |
Tab. č. xx: Přehled šestnácti faktorů monitorovaných Cattellovým osobnostním dotazníkem. U každého faktoru je uveden jeho název a příznačné charakteristiky osobnosti, v levém sloupci pro nízké skóre, v pravém sloupci pro vysoké skóre.
3.4. Průběh psychomotorického testování
Vliv latentní toxoplasmosy na výkon v psychomotorických testech byl zjišťován u dvou souborů osob: biologů a dárců krve.
Skupina biologů byla testována v standartním prostředí laboratoře. Kvůli zamezení případných diurnálních rozdílů ve výkonosti probíhala všechna vyšetření v dopoledních hodinách. Probandi byli tázani zda-li nejsou nachlazeni, neprodělali v poslední době infekční onemocnění a nekonzumovali alkoholické nápoje. V případě pozitivní odpovědi nebyli do výsledného souboru zahrnuti. Po uvedených dotazech a instrukcím k testům následovalo vlastní testování.
Stejně jako u souboru biologů probíhalo testování dárců krve v dopoledních hodinách v tiché místnosti. Z technických důvodů byl použit pouze test č.1. Všichni dárci podepsali před odběrem krve lékaři prohlášení, že neprodělali v posledních čtrnácti dnech žádné onemocnění a nekonzumovali předešlý večer alkoholické nápoje.
3.5. Psychomotorické testy
K hodnocení psychomotorické výkonosti byly použity dva počítačové testy užívané ve studiích vlivu virových infekcí na psychomotorický výkon (Smith a spol.1989; Hall & Smith 1994; Hall & Smith 1996). Oba testy měřily oko-motorickou koordinaci a udržení pozornosti.
Test č.1
V rozmezí 1-10 sec. se uprostřed monitoru objevoval podnět (čtverec). Proband byl instruován k co nejrychlejšímu zareagování při objevení podnětu zmáčknutím tlačítka. Tříminutový vlastní test, předcházela minutová zkouška. Byly vyhodnoceny průměrné reakční časy v jednotlivých minutách a celkový průměrný reakční čas.
Test č.2
V rychlém sledu (100 za minutu) se uprostřed monitoru objevovala trojciferná čísla, z nichž dvě po sobě následující se od sebe lišila pouze v jedné číslici (např. 397, 387, 478 atd.). Občas ( 8 krát za minutu) bezprostředně za sebou následovala stejná čísla. Úkolem probanda bylo detekovat tyto opakující se čísla a reagovat na ně, co nejrychleji, zmáčknutím tlačítka. Test opět tvořila minutová zkouška a tříminutový vlastní test. Hodnoceny byly po jednotlivých minutách a cekově počty správných a falešných reakcí a reakční čas správných reakcí.
3.6. Měření tělesné výšky, hmotnosti a výpočet indexu tělesné hmotnosti
Výška postavy byla měřena bez obuvi u stěny, v maximálním vzpřímení a s hlavou v přímé poloze (pohled jakoby do dálky) (Fetter 1967). K měření byl použit kalibrovaný antropometr s přesností 1 mm. K určování hmotnosti byla použita digitální pérová váha s přesností 0,5 kg. Měřený subjekt byl oblečen pouze v nejnutnějším spodním prádle. Index tělesné hmotnosti (BMI) byl vypočítán podle vzorce (Riegrová & Ulbrichová 1993):
BMI=hmotnost (kg)/(tělesná výška (m))2
3.7. Imunologické testy na toxoplasmosu
Specifická buněčná imunita byla v souboru biologů zjištěna intradermálním testem (Feldman 1954) v rámci předešlých studií (Flegr & Hrdý 1994; Flegr a spol. 1996). U souboru dárců krve a souboru pacientů byla existence specifických protilátek zjišťována metodou komplement fixační reakce (KFR) (Sabin & Waren 1942) v Národní referenční laboratoři pro toxoplasmosu při Státním zdravotním ústavu v Praze. Osoby byly na základě serologického vyšetření hodnoceny jako Toxoplasma pozitivní, pokud titry hladin protilátek byly vyšší nebo rovny 1:8.
3.8. Statistika
Na všechny statistické testy byl použit, není-li uvedeno jinak, program StatisticaŇ verze 5.0. Pro přehlednost jsou použité statistické metody uvedeny pro každou výsledkovou kapitolu zvlášť.
Analýza epidemiologických faktorů zvyšujících riziko infekce Toxoplasma gondii
Odpovědi nakažených a nenakažených osob na jednotlivé epidemiologické otázky byly zaznamenány do kontingenčních tabulek. Asociace každého epidemiologického faktoru a toxoplasmosy byla testována testem chi-kvadrát (Lepš 1996). Významnost efektu sledovaných faktorů a jejich vzájemných intetrakcí byla testována logistickou regresí (Hosmer & Lemeshow 1989). Tato metoda umožňuje pomocí logit transformace hodnotit vliv více poměrných i kategoriálních proměnných na závislou kategoriální proměnnou. Negativní odpovědi na jednotlivé epidemiogické otázky byly kódovány jako 0, pozitivní jako 1. Vzájemné dvojné interakce byly kódovány při pozitivní odpovědi na obě otázky jako 1, ostatní kombinace odpovědí jako 0 (Kleinbaum 1994). Výše rizika sledovaného faktoru byla odhadována pomocí Mantel-Haenszelova váženého odds ratio. Tato veličina byla vypočítána podle vzorce (Kahn & Sempeos 1989):
odds ratio=e b
kde e je základ přirozeného logaritmu a b standartizovaný regresní koeficient.
Konfidenční intervaly (95%) pro odds ratio byly vypočítány podle vzorce:
konf. int.=e b ± 1,96*SE
kde e je základ přirozeného logaritmu, b standartizovaný regresní koeficient a SE střední chyba odhadu b.
Analýza asociací faktorů osobnosti s epidemiologickými faktory zvyšujícími riziko infekce Toxoplasma gondii
Vliv osobnostních faktorů na sledovaný typ rizikového chování byl testován logistickou regresí (Hosmer & Lemeshow 1989). Jako nezávislé proměnné vstupovala hrubá skóre šestnácti osobnostních faktorů a matoucí proměnná věk, jako závislá proměnná sledovaný typ rizikového chování (chov kočky, konzumace syrového masa, chov králíků). Protože faktory osobnosti, v nichž byly zjištěny rozdíly mezi osobami nakaženými a nenakaženými, se lišily v závislosti na pohlaví, byla i zde hodnocena obě pohlaví zvlášť.
Změny osobnosti v důsledku latentní toxoplasmosy
K testování rozdílů v osobnostním profilu mezi nakaženými a nenakaženými osobami byla použita MANCOVA (Lepš 1996), kdy jako nezávislá proměnná vstupoval faktor toxoplasmosa, jako závislá proměnná hrubá skóre šestnácti osobnostních faktorů a jako kovariáta věk osoby. K statistickému hodnocení změn v osobnostním profilu mezi nakaženými a nenakaženými osobami byla opět použita analýza kovariance (ANCOVA) (Lepš 1996). Změny v jednotlivých faktorech byly vypočteny jako rozdíl mezi hrubým skórem dotazníku zasílaného po pěti letech a dotazníkem původním. Kladná hodnota tedy znamenala vzestup a vice versa. Protože změny v osobnostních faktorech nemusí mít lineární charakter, byl jako kovariáta použit věk a jeho druhá mocnina. Část osob dosahovala při původním testování v některých faktorech extrémních hodnot. V těchto případech již nebyla změna jedním směrem možná. Tento efekt byl odstraněn přidáním hrubého skóre v původním testu jako další kovariáty.
Vliv latentní toxoplasmosy na psychomotorický výkon
Shoda rozptylů byla ověřena Levenovým testem (anonymus 1995). Existence intersexuálních diferencí byla zjišťována t testem (Lepš 1996). V případě, že byl zjištěn rozdíl v rozptylech, byl efekt toxoplasmosy na psychomotorický výkon testován neparametrickým Kruskal-Wallis testem (anonymus 1995). Efekt věku byl při použití neparametrického testu odfiltrován použitím standartních residuálů z jednoduché lineární regrese mezi stářím osoby a příslušným reakčním časem. V ostatních případech byla použita jednocestná analýza kovariance (ANCOVA) (Lepš 1996), kdy jako kovariáta vstupovala proměnná věk. Protože u chronické infekce dochází k postupnému snižování hladin titrů protilátek, byly titry použity k odhadu délky infekce. Existence korelace mezi hladinou titrů protilátek a reakčním časem byla testována Kendallovým neparametrickým testem (anonymus 1995). Kvůli eliminaci efektu věku bylo použito místo reakčních časů standartních residuálů z jednoduché lineární regrese mezi stářím osoby a průměrným reakčním časem v příslušné minutě testu respektive v celém testu.
Prevalence toxoplasmosy u účastníků dopravních nehod
K statistickému hodnocení byl v tomto případě použit program EpiStatŇ verze 6. Na data bylo pohlíženo jako na věkově stratifikovanou studii případů a kontrol (age stratified case-control study) (Kahn & Sempeos 1989), kdy za případy byl považováni účastníci dopravních nehod a za kontroly výsledky serologických přehledů (Kodym a spol. 1995; Kodym a spol. 1997). Celkové vážené odds ratio bylo vypočítáno metodou maximálně pravděpodobnostního odhadu (maximum likelihood estimation) (Kahn & Sempeos 1989).
Vliv latentní toxoplasmozy na subjektivní hodnocení zdravotního stavu
Odpovědi na jednotlivé otázky byly stejně jako v analýze rizikových faktorů zaznamenány do kontingenčních tabulek a jejich asociace s toxoplasmosou testována Chi-kvadrát testem (Lepš 1996). Vliv latentní toxoplasmosy na subjektivní hodnocení zdravotního stavu s přihlédnutím ke stáří osoby byl testován logistickou regresí (Hosmer & Lemeshow 1989), kdy jako nezávislá binární proměnná vstupoval faktor toxoplasmosa a spojitá proměnná věk, jako závislá binární proměnná faktor zdravotní stav.
Vliv latentní toxoplasmosy na index tělesné hmotnosti
Shoda rozptylů byla ověřena Levenovým testem (anonymus 1995). Existence rozdílu v indexu tělesné hmotnosti (BMI) mezi skupinou nakažených a nenakažených osob byla testována jednocestnou analýzou kovariance (ANCOVA) (Lepš 1996), kdy jako nezávislá proměnná vstupoval faktor toxoplasmosa, jako závislá proměnná hodnota BMI a jako kovariáta věk. Analýza byla prováděna pro obě pohlaví zvlášť.
5. Výsledky
5.1. Analýza epidemiologických faktorů zvyšujících riziko infekce Toxoplasma gondii
Z celkového počtu 414 osob bylo na základě výsledků intradermálního testu 103 (24,9%) osob hodnoceno jako Toxoplasma pozitivních. Na epidemiologické dotazníky rozeslané 379 osobám jich odpovědělo celkem 334 (88,1 %) z toho 83 (24,9%) osob Toxoplasma pozitivních. Dotaz na velikost sídla v němž tázaná osoba prožila mládí byl položen v rámci Cattellova osobnostního dotazníku rozesílaného po pěti letech od původního vyplňování. Tento dotazník byl rozeslán celkem 373 osobám a odpovědělo na něj 268 osob.
Odpovědi Toxoplasma negativních a Toxoplasma pozitivních osob na jednotlivé otázky byly zaznamenány do kontingenčních tabulek. U otázky na frekvenci kontaktu s kočkou byla, kvůli malému počtu odpovědí v některých kategoriích, provedena redukce z původní šesti stupňové ordinální škály na škálu dvoustupňovou. Kategorie nazvaná zřídka vznikla spojením kategorií: nikdy, jednou za pět let a jednou za dva roky. Druhá kategorie nazvaná často vznikla spojením kategorií: jednou za rok, za měsíc a častěji. Mezi muži a ženami nebyl v žádném ze sledovaných faktorů zjištěn rozdíl a byla proto obě pohlaví hodnocena společně.
Analýzou kontingenčních tabulek byla zjištěna asociace mezi toxoplasmosou a chovem kočky (Chi-kvadrát=13,29, df=1, p=0,0003). Mezi osobami, které odpovídaly na otázku zda chovají (chovaly) kočku pozitivně, byla frekvence toxoplasmosy 32,5%, zatímco mezi osobami, které odpovídaly negativně, byla frekvence toxoplasmosy 15,1%. Četnosti odpovědí a relativní četnosti jsou uvedeny v tab. č. 5.
|
chov kočky |
|||
|
ne |
ano |
||
|
Toxoplasma negativní |
124 (84,9%) |
127 (67,5%) |
|
|
Toxoplasma pozitivní |
22 (15,1%) |
61 (32,5%) |
|
Tab. č. 5: Frekvenční tabulka pro faktory chov kočky a toxoplasmosa. V závorce uvedeny relativní četnosti ve sloupcích.
Na druhou stranu vztah mezi frekvencí styku s kočkou a toxoplasmosou zjištěn nebyl. Mezi osobami, které uváděly, že hladí kočku jednou za rok a častěji, byla frekvence Toxoplasma pozitivních 24,9%, zatímco mezi osobami uvádějícími hlazení kočky maximálně jednou za dva roky, byla frekvence Toxoplasma pozitivních 22,6%. Tento rozdíl je statisticky nesignifikantní (Chi-kvadrát=0,147, df=1, p=0,701). Četnosti a relativní četnosti odpovědí v původní formě jsou uvedeny v tab. č. 6A a v tab. č. 6B v redukované formě.
|
A |
kontakt s kočkou |
||||||
|
nikdy |
za pět let |
za dva roky |
za rok |
za měsíc |
častěji |
||
|
Toxoplasma negativní |
17 (70,8%) |
18 (85,7%) |
13 (76,5%) |
70 (77,8%) |
57 (83,8%) |
72 (67,3%) |
|
|
Toxoplasma pozitivní |
7 (29,2%) |
3 (14,3%) |
4 (23,5%) |
20 (22,2%) |
11 (16,2%) |
35 (32,7%) |
|
|
B |
kontakt s kočkou (reduk.) |
|
|
zřídka |
často |
|
|
Toxoplasma negativní |
48 (77,4%) |
199 (75,1%) |
|
Toxoplasma pozitivní |
14 (22,6%) |
66 (24,9%) |
Tab. č. 6: Frekvenční tabulka pro faktory kontakt s kočkou a toxoplasmosa. V závorce uvedeny relativní četnosti ve sloupcích. Tab. 6A původní forma a tab. 6B redukovaná forma.
Dále byla zjištěna statisticky významně vyšší frekvence Toxoplasma pozitivních osob u chovatelů králíků (32,9% oproti 17,1% u nechovatelů, Chi-kvadrát=11,255, df=1, p=0,0008). Četnosti a relativní četnosti odpovědí jsou uvedeny v tab. č. 7. Také konzumace syrového masa byla hodnocena jako významný rizikový faktor z hlediska získání toxolasmosy. U osob uvádějících konzumaci syrového masa byla frekvence Toxoplasma pozitivních 33,1%, kdežto mezi osobami, které konzumaci syrového masa neuváděly, byla frekvence Toxoplasma pozitivních 18,4% (Chi-kvadrát=9,533, df=1, p=0,002). Četnosti a relativní četnosti odpovědí jsou uvedeny v tab. č. 8.
|
chov králíků |
||
|
ne |
ano |
|
|
Toxoplasma negativní |
141 (82,9%) |
110 (67,1%) |
|
Toxoplasma pozitivní |
29 (17,1%) |
54 (32,9%) |
Tab. č. 7: Frekvenční tabulka pro faktory chov králíků a toxoplasmosa. V závorce uvedeny relativní četnosti ve sloupcích.
|
konzumace syrového masa |
||
|
ne |
ano |
|
|
Toxoplasma negativní |
151 (81,6%) |
99 (66,9%) |
|
Toxoplasma pozitivní |
34 (18,4%) |
49 (33,1%) |
Tab. č. 8: Frekvenční tabulka pro faktory konzumace syrového masa a toxoplasmosa. V závorce uvedeny relativní četnosti ve sloupcích.
Podle velikosti sídla, v němž tázaný subjekt prožil mládí, byla v kategorii obec do pěti tisíc obyvatel zjištěna frekvence Toxoplasma pozitivních osob 37,1%, v kategorii obec do 10000 obyvatel 28,2% a v kategorii nad 100000 obyvatel 16,8% Toxoplasma pozitivních osob (Chi-kvadrát=9,521, df=2, p=0,009). Za účelem analýzy interakcí jednotlivých epidemiologických faktorů bylo provedeno spojení kategorií do pěti tisíc obyvatel a do 100000 obyvatel. Touto redukcí vznikla dichotomická proměnná s kategoriemi do 100000 obyvatel a nad 100000 obyvatel. Frekvence nakažených osob v kategorii do 100000 obyvatel byla 31,9% oproti 16,8% v kategorii nad 100000 obyvatel. Zjištěná asociace s toxoplasmosou byla opět vysoce signifikantní (Chi-kvadrát=8,027, df=1, p=0,005). Četnosti a relativní četnosti odpovědí jsou uvedeny v tab. č. 9.
|
A |
velikost sídla (počet obyvatel) |
||
|
do 5000 |
do 100000 |
nad 100000 |
|
|
Toxoplasma negativní |
39 (62,9%) |
61 (71,8%) |
99 (83,2%) |
|
Toxoplasma pozitivní |
23 (37,1%) |
24 (28,2%) |
20 (16,8%) |
|
B |
velikost sídla (počet obyvatel) |
|
|
do 100000 |
nad 100000 |
|
|
Toxoplasma negativní |
100 (68,1%) |
99 (83,2%) |
|
Toxoplasma pozitivní |
47 (31,9%) |
20 (16,8%) |
Tab. č. 9: Frekvenční tabulka pro faktory velikost sídla a toxoplasmosa. V závorce uvedeny relativní četnosti ve sloupcích. Tab. 9A původní forma a tab. 9B redukovaná forma.
Chov psa zřejmě nehraje v epidemiologii toxoplasmosy významnou roli. Frekvence nakažených osob byla mezi chovateli psa 26,8% oproti 19,7% nakažených osob mezi nechovateli (Chi-kvadrát=2,516, df=1, p=0,113). Četnosti a relativní četnosti odpovědí jsou uvedeny v tab. č. 10.
|
chov psa |
||
|
ne |
ano |
|
|
Toxoplasma negativní |
100 (80,0%) |
151 (72,3%) |
|
Toxoplasma pozitivní |
25 (20,0%) |
58 (27,7%) |
Tab. č. 10: Frekvenční tabulka pro faktory chov psa a toxoplasmosa. V závorce uvedeny relativní četnosti ve sloupcích.
Analýzou kontingenčních tabulek byla zjištěna asociace mezi toxoplasmosou a těmito sledovanými faktory: chov kočky, konzumace syrového masa, chov králíků a velikost sídla. Některé asociace by však mohly být důsledkem vztahu s jiným sledovaným faktorem, který s toxoplasmosou přímo souvisí. Významnost jednotlivých rizikových faktorů a jejich vzájemných interakcí byla proto testována logistickou regresí. Do analýzy vstupovaly jako nezávislé binární proměnné: chov kočky, konzumace syrového masa, chov králíků, velikost sídla, všechny jejich vzájemné dvojné interakce (chov kočky-chov králíků, chov kočky-velikost sídla atd.) a rušivá proměnná věk, jako závislá proměnná vstupoval faktor toxoplasmosa. Celkový vysvětlovací efekt modelu byl vysoce signifikantní (Chi-kvadrát=39945, df=11, p=0,00004). Významný efekt vykazovaly faktory chov kočky (t(224)=2,203, p=0,029), konzumace syrového masa (t(224)=2,944 p=0,004) a interakce chov kočky-konzumace syrového masa (t(224)= -2,298, p=0,012), konzumace syrového masa-velikost sídla (t(224)= -1,869, p=0,021). Pokud byly z modelu odstraněny interakce u nichž nebyl zaznamenán významný efekt (chov kočky-chov králíků, chov kočky-velikost sídla, chov králíků-konzumace syrového masa, chov králíků-velikost sídla), nebyla vysvětlovací hodnota tohoto modelu (Chi-kvadrát=38,307, df=7, p=0,000003) horší oproti modelu původnímu (Chi-kvadrát=1,638, df=4, p=0,802). Významný efekt opět vykazovaly faktory chov kočky (t(228)=2,781, p=0,006), konzumace syrového masa (t(228)=3,751, p=0,0002), interakce chov kočky-konzumace syrového masa (t(228)=-2,53, p=0,012), konzumace syrového masa-velikost sídla (t(228)=-2,813, p=0,005) a v tomto případě byl významný i chov králíků (t(222)=2,243, p=0,026). Podle odhadu odds ratio zvyšuje chov kočky riziko získání toxoplasmosy 6,2 krát, konzumace syrového masa 19,4 krát a chov králíků 2,25 krát. Výsledky modelu jsou uvedeny v tab. č. 11.
|
konst.B0 |
věk |
chov kočky |
chov králíků |
konzumace syrového masa |
velikost sídla |
interakce kočka-maso |
interakce maso-sídlo |
|
|
odhad b |
-4,052 |
0,025 |
1,819 |
0,812 |
2,967 |
0,516 |
-2,074 |
-2,007 |
|
Odds ratio |
1,03 |
6,17 |
2,25 |
19,44 |
1,68 |
0,13 |
0,13 |
|
|
Konf. int. |
(1,00-1,05) |
(1,71-22,22) |
(1,11-4,58) |
(4,12-91,67) |
(0,64-4,41) |
(0,03-0,63) |
(0,03-0,54) |
|
|
SE |
0,807 |
0,014 |
0,654 |
0,362 |
0,791 |
0,494 |
0,819 |
0,713 |
|
t(228) |
-5,019 |
1,776 |
2,781 |
2,243 |
3,751 |
1,044 |
-2,530 |
-2,813 |
|
p |
0,000 |
0,077 |
0,006 |
0,026 |
0,0002 |
0,297 |
0,012 |
0,005 |
Tab. č. 11: Asociace toxoplasmosy s faktory chov kočky, chov králíků, konzumace syrového masa, interakcemi chov kočky-konzumace syrového masa, konzumace syrového masa-velikost sídla a matoucí proměnnou věk. V druhém sloupci je uveden odhad konstanty B0. Pro každý faktor je uveden standartizovaný regresní koeficient b, odds ratio, 95% konfideční intervaly pro odds ratio, střední chyba koeficientu b, hodnota t-testu a odpovídající hladina signifikance. Model logistická regrese.
5.2. Analýza asociací faktorů osobnosti s epidemiologickými faktory zvyšujícími riziko infekce Toxoplasma gondii
Do analýzy bylo zahrnuto 315 biologů (176 mužů a 139 žen), kteří odpověděli na epidemiologický dotazník a zároveň v rámci předešlých studií (Flegr & Hrdý 1994; Flegr a spol. 1996) vyplnili Cattellův šestnáctifaktorový osobnostní dotazník. Podle výsledků intradermálního testu bylo 240 osob (127 mužů a 113 žen) hodnoceno jako Toxoplasma negativní a 75 osob (49 mužů a 26 žen) jako Toxoplasma pozitivní. Analýzou interakcí mezi rizikovými faktory bylo zjištěno, že riziko infekce zvyšuje především chov kočky, konzumace syrového masa a chov králíků. Bylo proto testováno, zda-li těmto typům rizikového chování odpovídá určitý osobnostní profil, čili zda se v některých faktorech osobnosti liší např. chovatelé koček od nechovatelů.
Vliv osobnostních faktorů na sledovaný typ rizikového chování byl testován logistickou regresí. Jako nezávislá proměnná vstupovala hrubá skóre šestnácti osobnostních faktorů a matoucí proměnná věk, jako závislá proměnná sledovaný typ rizikového chování (chov kočky, konzumace syrového masa, chov králíků). Protože faktory osobnosti, v nichž byly zjištěny rozdíly mezi osobami nakaženými a nenakaženými, se lišily v závislosti na pohlaví, byla i zde hodnocena obě pohlaví zvlášť. Na základě výsledků logistické regrese byla ve skupině 176 mužů zjištěna asociace mezi chovem kočky a faktory osobnosti I (harria/premsia) (t(158)=-2,033, p=0,043), Q2 (závislost na skupině) (t(158)=-2,368, p=0,019) a Q3 (integrace osobnosti) (t(158)= -2,170, p=0,031). Muži chovatelé vykazovali nižší průměrné hrubé skóre jak ve faktorech I (tvrdý, realistický) a Q2 (závislý na skupině) tak i Q3 (špatně zformovaná osobnost) ve srovnání s muži, kteří kočku nechovají. Výsledky logistické regrese a průměrná hrubá skóre pro jednotlivé osobnostní faktory jsou uvedeny v tab. č. 12. Ve skupině 139 žen nebyla zjištěna žádná významná asociace mezi chovem kočky a faktory osobnosti. Významný byl v tomto případě efekt věku (t(121)=2,051, p=0,042). Výsledky logistické regrese a průměrná hrubá skóre pro jednotlivé osobnostní faktory jsou uvedeny v tab. č. 13.
Mezi faktorem konzumace syrového masa a osobnostními faktory nebyla zjištěna žádná významná korelace a to jak u mužů tak i u žen. Výsledky logistické regrese a průměrná hrubá skóre pro jednotlivé osobnostní faktory jsou uvedeny v tab. č. 14 pro muže a v tab. č. 15 pro ženy.
Chov králíků byl u mužů spojen s osobnostním faktorem Q2 (závislost na skupině) (t(158)=-3,099, p=0,002). Stejně jako v případě chovu kočky bylo i u mužů, kteří udávali, že chovají králíky zjištěno průměrné nižší hrubé skóre ve faktoru Q2 (závislý na skupině) ve srovnání s muži, kteří králíky nechovají. U žen je chov králíků spojen s průměrným nižším skórem ve faktorech B (inteligence) (t(121)=-2,302, p=0,023) a N (naivita) (t(121)=-2,577, p=0,011) Hranici signifikance se blížil i faktor A (afektotýmie) (t(121)=1,846, p=0,067). Výsledky logistické regrese a průměrná hrubá skóre pro jednotlivé osobnostní faktory jsou uvedeny v tab. č. 16 pro muže a v tab. č. 17 pro ženy.
|
chov kočky |
(průměr) |
odhad b |
SE |
t(158) |
p |
|
|
ne |
ano |
|||||
|
konst. B0 |
5,104 |
2,446 |
2,086 |
0,038 |
||
|
věk |
28,8 |
26,0 |
0,001 |
0,015 |
0,100 |
0,920 |
|
A |
9,78 |
9,51 |
0,017 |
0,063 |
0,282 |
0,778 |
|
B |
9,28 |
9,87 |
0,146 |
0,082 |
1,770 |
0,078 |
|
C |
13,83 |
13,86 |
0,024 |
0,061 |
0,406 |
0,684 |
|
E |
14,34 |
14,34 |
-0,027 |
0,052 |
-0,518 |
0,604 |
|
F |
11,52 |
11,85 |
-0,045 |
0,056 |
-0,811 |
0,418 |
|
G |
9,84 |
8,53 |
-0,029 |
0,055 |
-0,528 |
0,598 |
|
H |
11,10 |
10,14 |
-0,043 |
0,042 |
-1,034 |
0,302 |
|
I |
11,65 |
10,51 |
-0,099 |
0,048 |
-2,033 |
0,043 |
|
L |
10,48 |
10,58 |
0,00002 |
0,081 |
0,0001 |
0,999 |
|
M |
13,89 |
13,75 |
-0,036 |
0,059 |
-0,618 |
0,536 |
|
N |
8,15 |
7,67 |
-0,066 |
0,075 |
-0,873 |
0,383 |
|
O |
10,05 |
10,19 |
0,022 |
0,056 |
0,391 |
0,695 |
|
Q1 |
7,74 |
8,78 |
0,117 |
0,067 |
1,743 |
0,083 |
|
Q2 |
14,34 |
13,51 |
-0,154 |
0,065 |
-2,368 |
0,019 |
|
Q3 |
10,76 |
9,79 |
-0,147 |
0,067 |
-2,170 |
0,031 |
|
Q4 |
13,78 |
13,71 |
-0,038 |
0,051 |
-0,746 |
0,456 |
Tab č. 12: Korelace mezi chovem kočky a osobnostními faktory u mužů. První sloupec ukazuje průměrné hrubé skóre osobnostních faktorů podle toho zda osoby chovají, či nechovají kočku, v dalších sloupcích jsou uvedeny: standartizovaný regresní koeficient b, střední chyba koeficientu b, hodnota t-testu a dosažená hladina významnosti. V prvním řádku je uvedena konstanta B0. Model logistická regrese.
|
chov kočky (průměr) |
odhad b |
SE |
t(121) |
p |
|||
|
ne |
ano |
||||||
|
konst. B0 |
-0,528 |
2,672 |
-0,197 |
0,843 |
|||
|
věk |
20,9 |
23,0 |
0,081 |
0,039 |
2,051 |
0,042 |
|
|
A |
10,78 |
11,25 |
0,068 |
0,068 |
1,011 |
0,313 |
|
|
B |
9,59 |
9,41 |
-0,037 |
0,098 |
-0,386 |
0,699 |
|
|
C |
13,79 |
12,80 |
-0,042 |
0,073 |
-0,583 |
0,561 |
|
|
E |
12,83 |
11,46 |
-0,036 |
0,061 |
-0,607 |
0,544 |
|
|
F |
14,03 |
13,51 |
0,055 |
0,061 |
0,917 |
0,361 |
|
|
G |
9,203 |
9,78 |
0,019 |
0,059 |
0,329 |
0,742 |
|
|
H |
10,88 |
9,67 |
-0,044 |
0,052 |
-0,843 |
0,401 |
|
|
I |
14,00 |
14,44 |
-0,001 |
0,064 |
-0,015 |
0,987 |
|
|
L |
10,66 |
10,08 |
-0,071 |
0,064 |
-1,112 |
0,268 |
|
|
M |
13,12 |
12,82 |
-0,019 |
0,065 |
-0,299 |
0,765 |
|
|
N |
7,44 |
8,34 |
0,097 |
0,074 |
1,311 |
0,192 |
|
|
O |
11,81 |
12,58 |
0,006 |
0,064 |
0,106 |
0,915 |
|
|
Q1 |
7,31 |
7,33 |
0,044 |
0,066 |
0,672 |
0,502 |
|
|
Q2 |
12,95 |
12,55 |
-0,001 |
0,081 |
-0,016 |
0,987 |
|
|
Q3 |
10,46 |
9,95 |
-0,091 |
0,083 |
-1,088 |
0,278 |
|
|
Q4 |
14,68 |
15,52 |
-0,002 |
0,0621 |
-0,035 |
0,971 |
|
Tab č. 13: Korelace mezi chovem kočky a osobnostními faktory u žen. První sloupec ukazuje průměrné hrubé skóre osobnostních faktorů podle toho zda osoby chovají, či nechovají kočku, v dalších sloupcích jsou uvedeny: standartizovaný regresní koeficient b, střední chyba koeficientu b, hodnota t-testu a dosažená hladina významnosti. V prvním řádku je uvedena konstanta B0. Model logistická regrese.
|
konzumace syrového masa (průměr) |
odhad b |
SE |
t(158) |
p |
|||
|
ne |
ano |
||||||
|
konst. B0 |
-2,771 |
2,334 |
-1,186 |
0,237 |
|||
|
věk |
26,8 |
27,8 |
0,008 |
0,014 |
0,568 |
0,571 |
|
|
A |
9,63 |
9,62 |
-0,011 |
0,059 |
-0,192 |
0,847 |
|
|
B |
9,58 |
9,65 |
0,014 |
0,078 |
0,191 |
0,849 |
|
|
C |
13,91 |
13,76 |
0,033 |
0,057 |
0,585 |
0,558 |
|
|
E |
13,91 |
14,88 |
0,011 |
0,048 |
0,241 |
0,809 |
|
|
F |
11,57 |
11,87 |
0,012 |
0,052 |
0,243 |
0,807 |
|
|
G |
9,11 |
9,10 |
-0,017 |
0,053 |
-0,331 |
0,740 |
|
|
H |
10,05 |
11,20 |
0,047 |
0,039 |
1,206 |
0,229 |
|
|
I |
11,25 |
10,70 |
-0,045 |
0,043 |
-1,031 |
0,303 |
|
|
L |
10,35 |
10,76 |
-0,018 |
0,059 |
-0,318 |
0,751 |
|
|
M |
13,87 |
13,74 |
-0,009 |
0,056 |
-0,171 |
0,863 |
|
|
N |
7,99 |
7,74 |
-0,008 |
0,072 |
-0,122 |
0,902 |
|
|
O |
9,64 |
10,74 |
0,099 |
0,053 |
1,840 |
0,067 |
|
|
Q1 |
8,26 |
8,39 |
-0,014 |
0,062 |
-0,233 |
0,815 |
|
|
Q2 |
13,68 |
14,10 |
0,066 |
0,058 |
1,137 |
0,256 |
|
|
Q3 |
10,29 |
10,11 |
0,004 |
0,061 |
0,072 |
0,942 |
|
|
Q4 |
13,43 |
14,12 |
0,013 |
0,048 |
0,289 |
0,772 |
|
Tab č. 14: Korelace mezi faktorem konzumace syrového masa a osobnostními faktory u mužů. První sloupec ukazuje průměrné hrubé skóre osobnostních faktorů podle toho zda osoby konzumují, či nekonzumují syrové maso, v dalších sloupcích jsou uvedeny: standartizovaný regresní koeficient b, střední chyba koeficientu b, hodnota t-testu a dosažená hladina významnosti. V prvním řádku je uvedena konstanta BO. Model logistická regrese.
|
konzumace syrového masa (průměr) |
odhad b |
SE |
t(121) |
p |
|||
|
ne |
ano |
||||||
|
konst. B0 |
-3,382 |
2,739 |
-1,234 |
0,219 |
|||
|
věk |
21,2 |
23,1 |
0,063 |
0,032 |
1,938 |
0,054 |
|
|
A |
10,57 |
11,58 |
-0,021 |
0,069 |
-0,301 |
0,763 |
|
|
B |
9,39 |
9,56 |
0,061 |
0,101 |
0,599 |
0,550 |
|
|
C |
13,28 |
13,18 |
-0,091 |
0,077 |
-1,175 |
0,242 |
|
|
E |
11,21 |
12,87 |
0,068 |
0,061 |
1,103 |
0,272 |
|
|
F |
12,97 |
14,46 |
0,095 |
0,062 |
1,522 |
0,131 |
|
|
G |
9,32 |
9,87 |
0,026 |
0,062 |
0,427 |
0,669 |
|
|
H |
8,91 |
11,44 |
0,030 |
0,053 |
0,563 |
0,574 |
|
|
I |
14,09 |
14,46 |
0,054 |
0,066 |
0,831 |
0,407 |
|
|
L |
10,65 |
9,89 |
-0,112 |
0,066 |
-1,708 |
0,090 |
|
|
M |
12,86 |
13,04 |
0,026 |
0,066 |
0,401 |
0,688 |
|
|
N |
8,23 |
7,69 |
-0,091 |
0,074 |
-1,224 |
0,223 |
|
|
O |
12,28 |
12,27 |
0,072 |
0,066 |
1,078 |
0,283 |
|
|
Q1 |
7,28 |
7,38 |
-0,023 |
0,069 |
-0,341 |
0,733 |
|
|
Q2 |
13,13 |
12,29 |
-0,084 |
0,080 |
-1,053 |
0,294 |
|
|
Q3 |
10,09 |
10,30 |
0,093 |
0,083 |
1,123 |
0,263 |
|
|
Q4 |
15,36 |
14,89 |
0,013 |
0,063 |
0,206 |
0,836 |
|
Tab č. 15: Korelace mezi faktorem konzumace syrového masa a osobnostními faktory u žen. První sloupec ukazuje průměrné hrubé skóre osobnostních faktorů podle toho zda osoby konzumují, či nekonzumují syrové maso, v dalších sloupcích jsou uvedeny: standartizovaný regresní koeficient b, střední chyba koeficientu b, hodnota t-testu a dosažená hladina významnosti. V prvním řádku je uvedena konstanta B0. Model logistická regrese.
|
chov králíků (průměr) |
odhad b |
SE |
t(158) |
p |
|||
|
ne |
ano |
||||||
|
konst. B0 |
3,045 |
2,415 |
1,261 |
0,209 |
|||
|
věk |
27,5 |
26,9 |
0,004 |
0,015 |
0,273 |
0,784 |
|
|
A |
9,93 |
9,26 |
-0,091 |
0,061 |
-1,491 |
0,137 |
|
|
B |
9,47 |
9,77 |
0,072 |
0,081 |
0,894 |
0,372 |
|
|
C |
13,84 |
13,85 |
0,0007 |
0,058 |
0,012 |
0,990 |
|
|
E |
14,36 |
14,31 |
-0,007 |
0,049 |
-0,141 |
0,887 |
|
|
F |
11,39 |
12,07 |
0,031 |
0,053 |
0,579 |
0,563 |
|
|
G |
9,46 |
8,67 |
-0,025 |
0,054 |
-0,463 |
0,643 |
|
|
H |
10,89 |
10,16 |
-0,039 |
0,041 |
-0,965 |
0,335 |
|
|
I |
11,26 |
10,71 |
-0,046 |
0,045 |
-1,041 |
0,299 |
|
|
L |
10,30 |
10,82 |
0,074 |
0,062 |
1,199 |
0,232 |
|
|
M |
13,67 |
13,97 |
0,023 |
0,059 |
0,390 |
0,696 |
|
|
N |
7,82 |
7,95 |
0,067 |
0,074 |
0,896 |
0,371 |
|
|
O |
10,14 |
10,11 |
0,009 |
0,053 |
0,171 |
0,864 |
|
|
Q1 |
8,22 |
8,43 |
-0,011 |
0,063 |
-0,175 |
0,861 |
|
|
Q2 |
14,40 |
13,22 |
-0,196 |
0,063 |
-3,099 |
0,002 |
|
|
Q3 |
10,47 |
9,90 |
-0,054 |
0,064 |
-0,843 |
0,400 |
|
|
Q4 |
13,85 |
13,60 |
-0,046 |
0,049 |
-0,947 |
0,344 |
|
Tab č. 16: Korelace mezi faktorem chov králíků a osobnostními faktory u mužů. První sloupec ukazuje průměrné hrubé skóre osobnostních faktorů podle toho zda osoby chovají, či nechovají králíky, v dalších sloupcích jsou uvedeny: odhad standartizovaný regresní koeficient b, střední chyba koeficientu b, hodnota t-testu a dosažená hladina významnosti. V prvním řádku je uvedena konstanta B0. Model logistická regrese.
|
chov králíků (průměr) |
odhad b |
SE |
t(121) |
p |
|||
|
ne |
ano |
||||||
|
konst. B0 |
1,862 |
2,736 |
0,681 |
0,497 |
|||
|
věk |
21,7 |
22,4 |
0,037 |
0,031 |
1,176 |
0,241 |
|
|
A |
10,19 |
11,82 |
0,132 |
0,071 |
1,846 |
0,067 |
|
|
B |
9,95 |
9,06 |
-0,248 |
0,108 |
-2,302 |
0,023 |
|
|
C |
13,57 |
12,90 |
-0,065 |
0,074 |
-0,882 |
0,379 |
|
|
E |
11,63 |
12,41 |
0,048 |
0,062 |
0,768 |
0,443 |
|
|
F |
13,13 |
14,27 |
0,035 |
0,063 |
0,553 |
0,581 |
|
|
G |
9,60 |
9,47 |
-0,011 |
0,063 |
-0,170 |
0,865 |
|
|
H |
9,33 |
10,95 |
-0,019 |
0,053 |
-0,357 |
0,721 |
|
|
I |
14,25 |
14,24 |
0,017 |
0,065 |
0,266 |
0,791 |
|
|
L |
10,37 |
10,28 |
-0,078 |
0,067 |
-1,165 |
0,246 |
|
|
M |
12,98 |
12,91 |
-0,018 |
0,068 |
-0,269 |
0,788 |
|
|
N |
8,53 |
7,43 |
-0,210 |
0,081 |
-2,577 |
0,011 |
|
|
O |
12,03 |
12,46 |
0,0598 |
0,067 |
0,885 |
0,377 |
|
|
Q1 |
7,40 |
7,24 |
-0,048 |
0,070 |
-0,691 |
0,491 |
|
|
Q2 |
13,34 |
12,15 |
-0,067 |
0,082 |
-0,816 |
0,415 |
|
|
Q3 |
10,40 |
9,94 |
0,077 |
0,083 |
0,927 |
0,355 |
|
|
Q4 |
14,78 |
15,50 |
0,038 |
0,065 |
0,591 |
0,555 |
|
Tab č. 17: Korelace mezi faktorem chov králíků a osobnostními faktory u žen. První sloupec ukazuje průměrné hrubé skóre osobnostních faktorů podle toho zda osoby chovají, či nechovají králíky, v dalších sloupcích jsou uvedeny: standartizovaný regresní koeficient b, střední chyba koeficientu b, hodnota t-testu a dosažená hladina významnosti. V prvním řádku je uvedena konstanta B0. Model logistická regrese.
5.3. Změny osobnosti v důsledku latentní toxoplasmosy
Soubor dárců krve
Osobnostní dotazník byl předán celkem 101 osobě. Vyplněný dotazník zaslalo zpět 57 osob (56,4%), ve dvou případech byl dotazník vyplněn nekompletně a nemohl být proto použit. Analýza byla tedy provedena na základě údajů získaných od 24 žen (z toho 11 nenakažených) a 31 mužů (z toho 19 nenakažených). Rozdíly v psychologickém profilu mezi skupinou Toxoplasma negativních a Toxoplasma pozitivních osob byly testovány analýzou kovariance (ANCOVA), přičemž věk osoby byl použit jako kovariáta. Protože je známo, že změny v některých osobnostních faktorech mají opačný směr u mužů a u žen (Flegr a spol. 1996), byly i zde obě pohlaví testována odděleně. Na základě výsledků předešlých studií (Flegr, Hrdý 1994; Flegr a spol. 1996) byly změny v určitých faktorech osobnosti očekávány, byl tedy použit jednostranný test namísto dvoustranného.
Mezi skupinou nakažených a nenakažených osob byl u mužů zjištěn významný rozdíl v osobnostních faktorech G (F(1,28)=3,198, p=0,043, jednostranný test) a N (F(1,28)=4,197, p=0,050, dvoustranný test). Toxoplasma pozitivní muži dosahovali oproti Toxoplasma negativním mužům nižšího skóre jak ve faktoru G (nízká síla superega), tak i N (naivita). Průměrná hrubá skóre jednotlivých osobnostních faktorů jsou uvedena v tab. č. 18.
V souboru žen vykazovaly rozdíly mezi nakaženými a nenakaženými osobami faktory A (F(1,21)=6,006, p=0,013, jednostranný test) a N (F(1,21)=4,77, p=0,040, dvoustranný test). Infekce toxoplasmou je tedy u žen spojena s vyšším faktorem A (afektothýmie) a narozdíl od mužů s vyšším faktorem N (naivita). Průměrná hrubá skóre jednotlivých osobnostních faktorů jsou uvedena v tab. č. 19.
|
Toxoplasma |
negativní |
Toxoplasma |
pozitivní |
F |
p |
|
|
průměr |
sm.odch. |
průměr |
sm.odch. |
|||
|
A |
11,05 |
3,22 |
9,67 |
3,52 |
0,909 |
0,349 |
|
B |
8,73 |
2,18 |
7,91 |
1,44 |
0,925 |
0,344 |
|
C |
14,73 |
4,79 |
15,5 |
4,73 |
0,289 |
0,595 |
|
E |
13,26 |
3,38 |
12,75 |
4,63 |
0,307 |
0,584 |
|
F |
9,78 |
3,29 |
9,91 |
3,94 |
0,077 |
0,783 |
|
G |
12,84 |
3,92 |
10,33 |
3,20 |
3,198 |
0,043 * |
|
H |
9,94 |
5,55 |
11,41 |
4,29 |
0,586 |
0,450 |
|
I |
7,47 |
2,93 |
8,75 |
4,35 |
0,993 |
0,328 |
|
L |
9,15 |
3,77 |
10,08 |
3,17 |
0,384 |
0,270 * |
|
M |
12,94 |
3,67 |
12,5 |
2,54 |
0,222 |
0,641 |
|
N |
11,21 |
3,13 |
8,92 |
2,81 |
4,197 |
0,050 |
|
O |
9,21 |
3,44 |
7,92 |
4,12 |
0,869 |
0,180 * |
|
Q1 |
5,63 |
2,54 |
6,42 |
2,81 |
0,931 |
0,343 |
|
Q2 |
12,52 |
3,41 |
12,58 |
3,31 |
0,001 |
0,490 * |
|
Q3 |
13,84 |
3,38 |
12,41 |
2,67 |
1,621 |
0,213 |
|
Q4 |
11,73 |
5,03 |
12,33 |
4,62 |
0,046 |
0,831 |
Tab. č. 18: Průměrná hrubá skóre jednotlivých faktorů osobnosti monitorovaných Cattellovým dotazníkem. u mužů. * označeny faktory u nichž byl použit jednostranný test. MANCOVA.
|
Toxoplasma |
negativní |
Toxoplasma |
pozitivní |
F |
p |
|
|
průměr |
sm.odch. |
průměr |
sm.odch. |
|||
|
A |
9,72 |
2,72 |
12,46 |
2,66 |
6,006 |
0,012 * |
|
B |
8,54 |
2,42 |
8,76 |
1,58 |
0,051 |
0,823 |
|
C |
14,36 |
3,17 |
14,38 |
3,47 |
0,003 |
0,959 |
|
E |
12,0 |
5,58 |
14,15 |
3,51 |
1,175 |
0,291 |
|
F |
12,36 |
5,04 |
13,0 |
4,16 |
0,105 |
0,749 |
|
G |
11,81 |
4,06 |
11,53 |
4,13 |
0,099 |
0,757 |
|
H |
11,09 |
7,41 |
12,69 |
5,64 |
0,330 |
0,572 |
|
I |
13,27 |
3,37 |
12,76 |
3,26 |
0,246 |
0,625 |
|
L |
8,91 |
4,61 |
10,31 |
3,77 |
0,599 |
0,224 * |
|
M |
11,73 |
2,53 |
13,07 |
2,84 |
1,325 |
0,263 |
|
N |
9,72 |
2,72 |
12,23 |
2,61 |
4,770 |
0,040 |
|
O |
12,27 |
3,66 |
10,46 |
3,68 |
1,816 |
0,095 * |
|
Q1 |
6,0 |
1,73 |
6,76 |
2,38 |
0,832 |
0,372 |
|
Q2 |
11,73 |
4,07 |
13,38 |
3,52 |
0,983 |
0,166 * |
|
Q3 |
12,0 |
2,93 |
12,07 |
3,14 |
0,001 |
0,972 |
|
Q4 |
14,18 |
3,68 |
14,07 |
4,57 |
0,015 |
0,905 |
Tab. č. 19: Průměrná hrubá skóre jednotlivých faktorů osobnosti monitorovaných Cattellovým dotazníkem. u žen. * označeny faktory u nichž byl použit jednostranný test. MANCOVA.
Soubor biologů
Dotazník byl zaslán po pěti letech od původního testování 218 mužům a 155 ženám. Na základě výsledků intradermálního testu bylo 59 mužů (27,1%) a 31 žen (20%) hodnoceno jako Toxoplasma pozitivní.Vyplněný dotazník byl získán od 102 nenakažených a 42 (41,2%) nakažených mužů a od 100 nenakažených a 24 (24%) nakažených žen. U mužů byl zaznamenán významný rozdíl (p=0,009) ve frekvenci infekce mezi respondenty (42,1%) a nerespondenty (23%). Znamená to tedy, že nakažení muži odpovídali na dotazník relativně častěji než muži nenakažení. U žen podobný efekt efekt zaznamenán nebyl.
V první části analýzy byly zjišťováno zda je možné zaznamenat rozdíly v psychologickém profilu mezi nakaženými a nenakaženými osobami také na základě dotazníků zasílaných po pěti letech od původního testování. Protože změny v některých faktorech osobnosti zjištěné v předešlých studiích (Flegr, Hrdý 1994; Flegr a spol. 1996) měly opačný směr u mužů a u žen, byla i zde obě pohlaví testována odděleně. K testování byla použita analýza kovariance (ANCOVA), kdy jako nezávislá proměnná vstupoval faktor toxoplasmosa, jako závislá proměnná hrubé skóre jednotlivých faktorů osobnosti a jako kovariáta věk .
V souboru mužů byl zjištěn významný rozdíl mezi osobami Toxoplasma negativními a Toxoplasma pozitivními osobami ve faktorech B (F(1,141)=4,896, p=0,029) a L (F(1,141)=4,015, p=0,024, jednostranný test). Nakažení muži dosahovali ve srovnání s nenakaženými vyššího skóre ve faktoru B (inteligence) a stejně jako při původním testování vyššího skóre ve faktoru L (protenze). Mezi Toxoplasma negativními a Toxoplasma pozitivními ženami nebyl zjištěn rozdíl v žádném z šestnácti faktorů osobnosti. Průměrná hrubá skóre jednotlivých osobnostních faktorů jsou uvedena v tab. č. 20 pro muže a v tab. č. 21 pro ženy.
V další části analýzy bylo testováno zda dochází v důsledku infekce k systematickým změnám v osobnostním profilu. Změny v jednotlivých faktorech byly vypočteny jako rozdíl mezi hrubým skórem dotazníku zasílaného po pěti letech a dotazníkem původním. Kladná hodnota tedy znamenala vzestup a vice versa. K statistickému hodnocení byla opět použita analýza kovariance (ANCOVA). Protože změny v osobnostních faktorech nemusí mít lineární charakter, byl jako kovariáta použit věk a jeho druhá mocnina. U všech sledovaných osobnostních faktorů bylo zjištěno, že směr změny negativně koreluje s hodnotou faktoru zjištěného před pěti lety (viz graf č. 1 a 2). Tento efekt byl odstraněn přidáním hrubého skóre v původním testu jako další kovariáty.
V souboru mužů byly mezi nakaženými a nenakaženými osobami zaznamenány významné rozdíly ve změnách faktorů B (inteligence) (F(1,139)=7,536, p=0,007) a Q1 (radikalismus) (F(1,139)=4,713, p=0,032). U faktoru B došlo k průměrnému zvýšení u nakažených mužů ve srovnání s nenakaženými. U faktoru Q1 došlo k průměrnému snížení u nenakažených mužů ve srovnání s nakaženými. V souboru žen byly mezi nakaženými a nenakaženými osobami zjištěny rozdíly ve změnách faktoru Q4 (ergická tenze) (F(1,119)=8,58, p=0,004). Zatímco u Toxoplasma negativních žen došlo po pěti letech k poklesu tohoto faktoru, naopak u Toxoplasma pozitivních žen k jeho vzestupu. Průměrné změny v jednotlivých osobnostních faktorech jsou uvedeny v tab. č. 22 pro muže a v tab. č. 23 pro ženy. Nicméně pokud byla provedena Bonfferroniho korekce pro mnohonásobná porovnávání došlo u všech faktorů k poklesu signifikance testu pod hladinu 5%.
Výsledky analýzy změn osobnostních faktorů po pěti letech mohla ovlivnit nízká stabilita jednotlivých faktorů osobnosti. Stabilitou se rozumí, jednak nezávislost měření na náhodném kolísání v čase, jednak stálost vlastnosti samé (Říčan 1975). Byla proto zjišťována korelace mezi hodnotou původního faktoru a hodnotou faktoru zjištěného po pěti letech. Výsledné korelační koeficienty byly srovnány se standarty pro Cattellův dotazník, zjištěnými retestem po čtyřech letech (Cattell 1970). Ve všech případech byly zjištěné korelační koeficienty vyšší nebo se jen nevýznamně lišily od hodnot udávaných Cattellem. Přehled korelačních koeficientů pro jednotlivé faktory je uveden v tab. č. 24.

Graf č. 1: Korelace mezi skórem zjištěným při původním testování (L) a změnou po pěti letech (L1-L) v osobnostním faktoru L u mužů.
Graf č. 2: Korelace mezi skórem zjištěným při původním testování (L) a změnou po pěti letech (L1-L) v osobnostním faktoru L u žen.
|
Toxoplasma |
negativní |
Toxoplasma |
pozitivní |
F |
p |
|
|
průměr |
sm.odch. |
průměr |
sm.odch. |
|||
|
A |
9,44 |
3,19 |
8,78 |
3,45 |
1,114 |
0,293 |
|
B |
10,05 |
2,04 |
10,71 |
1,55 |
4,896 |
0,029 |
|
C |
14,58 |
3,72 |
13,38 |
4,57 |
2,780 |
0,098 |
|
E |
14,76 |
4,69 |
15,54 |
4,08 |
1,219 |
0,271 |
|
F |
11,85 |
4,79 |
11,57 |
4,84 |
0,001 |
0,973 |
|
G |
10,08 |
4,36 |
9,59 |
3,52 |
1,016 |
0,158 * |
|
H |
12,08 |
6,01 |
11,92 |
5,91 |
0,054 |
0,817 |
|
I |
10,69 |
3,42 |
9,92 |
3,71 |
1,457 |
0,229 |
|
L |
10,14 |
3,17 |
11,31 |
3,63 |
4,015 |
0,024 * |
|
M |
14,24 |
2,85 |
14,47 |
3,12 |
0,103 |
0,749 |
|
N |
8,02 |
2,96 |
7,83 |
2,41 |
0,261 |
0,610 |
|
O |
9,54 |
4,08 |
10,33 |
4,39 |
0,996 |
0,160 * |
|
Q1 |
7,34 |
2,68 |
8,42 |
2,85 |
5,586 |
0,019 |
|
Q2 |
13,84 |
3,24 |
14,12 |
2,73 |
0,165 |
0,343 * |
|
Q3 |
11,71 |
3,41 |
10,76 |
2,87 |
3,043 |
0,083 |
|
Q4 |
13,07 |
4,57 |
13,69 |
4,83 |
0,602 |
0,439 |
Tab. č. 20: Průměrná hrubá skóre jednotlivých faktorů osobnosti monitorovaných Cattellovým dotazníkem u mužů. * označeny faktory u nichž byl použit jednostranný test. ANCOVA.
|
Toxoplasma |
negativní |
Toxoplasma |
pozitivní |
F |
p |
|
|
průměr |
sm.odch. |
průměr |
sm.odch. |
|||
|
A |
10,53 |
3,51 |
10,92 |
3,69 |
0,508 |
0,235 * |
|
B |
10,47 |
1,81 |
10,41 |
1,86 |
0,007 |
0,933 |
|
C |
13,33 |
3,67 |
13,75 |
3,40 |
0,105 |
0,746 |
|
E |
12,69 |
4,06 |
12,17 |
4,82 |
0,625 |
0,431 |
|
F |
12,67 |
4,46 |
12,42 |
3,71 |
0,024 |
0,878 |
|
G |
10,82 |
4,07 |
11,67 |
4,25 |
0,328 |
0,568 |
|
H |
10,30 |
5,56 |
12,17 |
6,60 |
1,066 |
0,304 |
|
I |
13,59 |
3,31 |
13,17 |
3,34 |
0,596 |
0,442 |
|
L |
9,63 |
3,32 |
9,04 |
3,45 |
0,359 |
0,275 * |
|
M |
13,42 |
3,09 |
13,0 |
2,99 |
0,330 |
0,567 |
|
N |
8,25 |
2,60 |
9,04 |
3,54 |
1,732 |
0,191 |
|
O |
11,48 |
4,10 |
11,37 |
4,26 |
0,175 |
0,338 * |
|
Q1 |
6,49 |
2,84 |
6,67 |
2,68 |
0,122 |
0,728 |
|
Q2 |
13,24 |
3,54 |
12,79 |
3,52 |
0,324 |
0,285 * |
|
Q3 |
11,52 |
3,55 |
11,5 |
2,65 |
0,061 |
0,805 |
|
Q4 |
14,21 |
4,61 |
15,08 |
4,44 |
1,451 |
0,231 |
Tab. č. 21: Průměrná hrubá skóre jednotlivých faktorů osobnosti monitorovaných Cattellovým dotazníkem u žen. * označeny faktory u nichž byl použit jednostranný test. ANCOVA.
|
Toxoplasma |
negativní |
Toxoplasma |
pozitivní |
F |
p |
|
|
průměr |
sm.odch. |
průměr |
sm.odch. |
|||
|
A1-A |
-0,39 |
3,34 |
-0,64 |
2,61 |
0,698 |
0,404 |
|
B1-B |
0,34 |
2,11 |
1,21 |
2,06 |
7,536 |
0,007 |
|
C1-C |
0,51 |
3,18 |
-0,04 |
3,27 |
2,367 |
0,126 |
|
E1-E |
0,84 |
3,91 |
0,83 |
3,88 |
0,448 |
0,504 |
|
F1-F |
0,21 |
3,81 |
-0,02 |
3,51 |
0,071 |
0,791 |
|
G1-G |
0,36 |
3,52 |
0,83 |
2,93 |
0,027 |
0,869 |
|
H1-H |
1,37 |
4,72 |
0,95 |
4,51 |
0,212 |
0,645 |
|
I1-I |
-0,55 |
3,47 |
-0,71 |
3,37 |
0,760 |
0,384 |
|
L1-L |
-0,11 |
3,30 |
-0,07 |
3,40 |
1,502 |
0,222 |
|
M1-M |
0,29 |
3,59 |
0,64 |
3,17 |
0,181 |
0,670 |
|
N1-N |
-0,11 |
3,06 |
0,09 |
2,12 |
0,008 |
0,929 |
|
O1-O |
-0,28 |
3,49 |
-0,62 |
3,49 |
0,007 |
0,931 |
|
Q11-Q1 |
-0,63 |
3,31 |
0,05 |
2,81 |
4,713 |
0,031 |
|
Q21-Q2 |
0,205 |
3,28 |
0,19 |
2,54 |
0,023 |
0,879 |
|
Q31-Q3 |
1,06 |
3,03 |
0,57 |
2,91 |
2,01 |
0,159 |
|
Q41-Q4 |
-0,47 |
3,85 |
-0,33 |
3,79 |
0,301 |
0,583 |
Tab. č. 22: Průměrné změny v hrubém skóre jednotlivých faktorů osobnosti monitorovaných Cattellovým dotazníkem u mužů. ANCOVA.
|
Toxoplasma |
negativní |
Toxoplasma |
pozitivní |
F |
p |
|
|
průměr |
sm.odch. |
průměr |
sm.odch. |
|||
|
A1-A |
0,20 |
3,13 |
-1,67 |
2,68 |
1,28 |
0,259 |
|
B1-B |
0,98 |
2,07 |
0,87 |
1,75 |
0,037 |
0,846 |
|
C1-C |
0,36 |
2,77 |
-0,12 |
3,98 |
0,129 |
0,719 |
|
E1-E |
1,06 |
4,01 |
-1,08 |
5,31 |
3,50 |
0,063 |
|
F1-F |
-0,74 |
4,29 |
-1,167 |
3,93 |
0,307 |
0,580 |
|
G1-G |
1,13 |
3,28 |
0,91 |
3,41 |
0,167 |
0,683 |
|
H1-H |
0,95 |
4,23 |
-0,04 |
3,39 |
0,486 |
0,486 |
|
I1-I |
-0,73 |
2,83 |
-0,25 |
3,08 |
0,101 |
0,750 |
|
L1-L |
-0,70 |
3,96 |
-0,71 |
3,27 |
0,081 |
0,776 |
|
M1-M |
0,58 |
3,72 |
0,75 |
3,37 |
0,098 |
0,754 |
|
N1-N |
-0,01 |
2,92 |
0,75 |
2,95 |
2,34 |
0,128 |
|
O1-O |
-0,68 |
3,83 |
-0,21 |
3,67 |
0,304 |
0,582 |
|
Q11-Q1 |
-1,01 |
3,11 |
0,25 |
3,02 |
1,65 |
0,201 |
|
Q21-Q2 |
0,40 |
3,03 |
0,00 |
3,09 |
0,114 |
0,735 |
|
Q31-Q3 |
1,11 |
3,43 |
0,96 |
3,00 |
0,00006 |
0,994 |
|
Q41-Q4 |
-1,15 |
4,09 |
1,79 |
3,29 |
8,580 |
0,004 |
Tab. č. 23: Průměrné změny v hrubém skóre jednotlivých faktorů osobnosti monitorovaných Cattelovým dotazníkem u žen. ANCOVA.
|
muži |
ženy |
|||||||
|
celkem |
T. neg. |
T. pozit. |
Cattell |
celkem |
T. neg. |
T. pozit. |
Cattell |
|
|
A |
0,57 |
0,48 |
0,74 |
0,49 |
0,61 |
0,61 |
0,70 |
0,62 |
|
B |
0,45 |
0,47 |
0,47 |
0,28 |
0,52 |
0,53 |
0,50 |
0,23 |
|
C |
0,67 |
0,65 |
0,71 |
0,45 |
0,67 |
0,72 |
0,43 |
0,48 |
|
E |
0,65 |
0,67 |
0,60 |
0,47 |
0,53 |
0,59 |
0,41 |
0,52 |
|
F |
0,68 |
0,65 |
0,73 |
0,48 |
0,59 |
0,58 |
0,65 |
0,52 |
|
G |
0,66 |
0,65 |
0,69 |
0,54 |
0,68 |
0,68 |
0,63 |
0,46 |
|
H |
0,68 |
0,66 |
0,72 |
0,49 |
0,76 |
0,72 |
0,88 |
0,64 |
|
I |
0,57 |
0,51 |
0,67 |
0,63 |
0,63 |
0,63 |
0,60 |
0,42 |
|
L |
0,49 |
0,45 |
0,53 |
0,40 |
0,39 |
0,38 |
0,46 |
0,42 |
|
M |
0,36 |
0,37 |
0,37 |
0,43 |
0,32 |
0,30 |
0,40* |
0,49 |
|
N |
0,47 |
0,41 |
0,64 |
0,39 |
0,51 |
0,46 |
0,66 |
0,21 |
|
O |
0,65 |
0,64 |
0,65 |
0,57 |
0,57 |
0,55 |
0,64 |
0,52 |
|
Q1 |
0,41 |
0,36 |
0,52 |
0,52 |
0,47 |
0,51 |
0,32* |
0,51 |
|
Q2 |
0,49 |
0,46 |
0,60 |
0,46 |
0,60 |
0,61 |
0,55 |
0,50 |
|
Q3 |
0,55 |
0,55 |
0,55 |
0,41 |
0,54 |
0,54 |
0,51 |
0,41 |
|
Q4 |
0,65 |
0,64 |
0,67 |
0,56 |
0,62 |
0,63 |
0,72 |
0,51 |
Tab. č. 24: Přehled korelačních koeficientů (test-retest 5 let) pro jednotlivé faktory vyjadřujících míru jejich stability. Pro srovnání jsou uvedeny standarty pro Cattellův dotazník zjištěné retestem po 4 letech (Cattell 1970). * označeny korelační koeficienty nesignifikantní na hladině 5%.
5.4.Vliv latentní toxoplasmosy na psychomotorický výkon
Soubor biologů
Podle výsledků intradermálního testu na toxoplasmosu bylo 38 osob (23 mužů a 15 žen) hodnoceno jako pozitivní a 79 osob(38 mužů a 41 žen) jako negativní.
Test č.1
Z důvodu zabránění vlivu odlehlých hodnot byl u každé osoby vypočítán celkový průměrný reakční čas a jednotlivé reakční časy přesahující tuto hodnotu o 2,5 směrodatné odchylky byly z další analýzy vyloučeny. Mezi skupinou mužů a žen nebyl zjištěn rozdíl v reakčním čase a byly proto dále obě pohlaví hodnocena společně. Protože v první minutě testu a celkovém hodnocení byl zjištěn mezi skupinou nakažených a nenakažených osob rozdíl ve variancích (F=11,322, p=0,001, respektive F=5,032, p=0,027), byl k hodnocení reakčních časů použit neparametrický Kruskal-Wallis test. Efekt věku byl v těchto případech eliminován použitím standartních residuálů z jednoduché lineární regrese mezi reakčním časem a věkem. V 2. a 3. minutě testu byla k hodnocení použita jednocestná analýza kovariance (ANCOVA), do níž jako nezávislá proměnná vstupoval faktor toxoplasmosa, jako závislá proměnná reakční čas v druhé respektive třetí minutě testu a jako kovariáta věk. Průměrné reakční časy v jednotlivých minutách a i celkový průměrný reakční čas byly nižší u osob Toxoplasma negativních oproti osobám Toxoplasma pozitivních. Nicméně tyto rozdíly byly ve všech případech nesignifikantní. Výsledky jednotlivých testů a průměrné hodnoty reakčních časů jsou uvedeny v tab. č. 25. Průměrné reakční časy nakažených a nenakažených osob jsou zobrazeny v grafu č. 3.
|
reakční čas (milisec.) |
||||||||||
|
1. minuta |
2. minuta |
3. minuta |
1.-3. minuta |
|||||||
|
Toxoplasma negativní |
průměr |
246,3 |
252,6 |
255,4 |
251,4 |
|||||
|
sm. odch. |
22,8 |
24,9 |
30,4 |
23,1 |
||||||
|
Toxoplasma pozitivní |
průměr |
258,3 |
255,3 |
265,6 |
260,0 |
|||||
|
sm. odch. |
36,4 |
28,0 |
32,0 |
29,6 |
||||||
|
F (H) |
(0,693) |
0,177 |
2,091 |
(0,673) |
||||||
|
df (N) |
(117) |
115 |
115 |
(117) |
||||||
|
p |
0,405 |
0,675 |
0,151 |
0,412 |
||||||
Tab. č.25: Průměrné reakční časy (v milisec.) v jednotlivých minutách a celkový průměrný reakční čas. 1. minuta a 1.-3. minuta Kruskal-Wallis test, 2. minuta a 3. minuta ANCOVA.

Graf č. 3: Průměrné reakční časy (v milisec.) v jednotlivých minutách a celkově. Chybové úsečky označují směrodatnou odchylku, světlé sloupce osoby Toxoplasma negativní a tmavé sloupce osoby Toxoplasma pozitivní.
Test č.2
V tomto testu byl hodnocen počet zaregistrovaných podnětů (zásah) a počet reakcí jimž podnět nepředcházel (falešná reakce). U reakcí jimž předcházel podnět byla hodnocena i latence odpovědi (reakční čas). Vyhodnoceny byly reakce v jednotlivých minutách a celkově. Stejně jako v případě prvního testu nebyl mezi muži a ženami zjištěn rozdíl (hodnoceno t testem) ani v jedné ze sledovaných veličin. Další analýza byla proto prováděna pro obě pohlaví společně. Ve všech sledovaných minutách osoby Toxoplasma negativní reagovaly na podnět častěji, rychleji (kromě 3. minuty) a vykazovaly měně falešných reakcí oproti osobám Toxoplasma pozitivním. V žádném případě však tyto rozdíly nebyly signifikantní (ANCOVA, kovariáta věk). Naměřené hodnoty jsou uvedeny v tab. č. 26. a zobrazeny v grafech č. 4, 5 a 6.
|
Toxoplasma negativní |
Toxoplasma pozitivní |
F |
df |
p |
||
|
1.minuta |
reakční čas (milisec.) |
554,8 (95,2) |
581,4 (106,6) |
1,62 |
113 |
0,206 |
|
počet zásahů |
5,5 (1,6) |
4,8 (1,8) |
2,122 |
114 |
0,148 |
|
|
počet falešných reakcí |
4,5 (2,9) |
4,9 (4,9) |
0,609 |
114 |
0,437 |
|
|
2.minuta |
reakční čas (milisec.) |
583,1 (108,5) |
616,2 (111,6) |
1,802 |
112 |
0,182 |
|
počet zásahů |
4,8 (1,8) |
4,6 (1,6) |
0,785 |
114 |
0,377 |
|
|
počet falešných reakcí |
3,4 (2,3) |
3,7 (2,1) |
0,117 |
114 |
0,733 |
|
|
3.minuta |
reakční čas (milisec.) |
658,6 (143,1) |
649,5 (117,3) |
0,192 |
111 |
0,662 |
|
počet zásahů |
3,8 (1,6) |
3,4 (1,6) |
0,852 |
114 |
0,358 |
|
|
počet falešných reakcí |
3,8 (2,5) |
4,2 (2,1) |
1,254 |
114 |
0,265 |
|
|
1.-3. minuta |
reakční čas (milisec.) |
593,5 (90,9) |
612,5 (87,3) |
0,833 |
114 |
0,363 |
|
počet zásahů |
14,0 (3,77) |
12,8 (3,75) |
2,064 |
114 |
0,154 |
|
|
počet falešných reakcí |
11,6 (6,0) |
12,9 (7,8) |
0,895 |
114 |
0,346 |
Tab. č.26: Reakční čas, počet zásahů a falešných reakcí v jednotlivých minutách a celkově. V závorkách uvedeny směrodatné odchylky. ANCOVA.
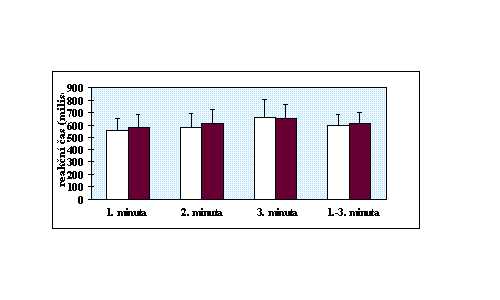
Graf č. 4: Průměrné reakční časy (v milisec.)
v jednotlivých minutách a celkově. Chybové úsečky označují směrodatnou
odchylku, světlé sloupce osoby nenakažené a tmavé sloupce osoby nakažené.
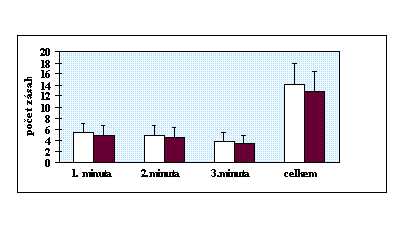
Graf č.5: Průměrné počty zásahů v jednotlivých minutách a během celého testu. Chybové úsečky označují směrodatnou odchylku, světlé sloupce osoby nenakažené a tmavé sloupce osoby nakažené.
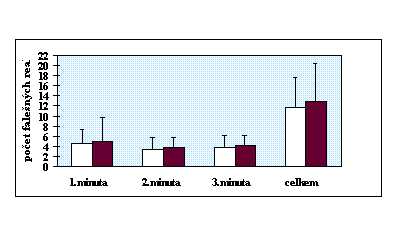
Graf č. 6: Průměrný počet falešných reakcí v jednotlivých minutách a v rámci celého testu. Chybové úsečky označují směrodatnou odchylku, světlé sloupce osoby nenakažené a tmavé sloupce osoby nakažené.
Soubor dárců krve
Z celkem 117 testovaných osob bylo na základě serologického vyšetření testem KFR 61 osob (33 mužů a 28 žen) hodnoceno jako Toxoplasma pozitivní. U jedné osoby však svědčila hladina protilátek na nedávno prodělanou akutní toxoplasmosu (titr 1:2048) a byla proto z další analýzy vyloučena.
Test č.1
Stejně jako v souboru biologů nebyly do analýzy zahrnuty odlehlé hodnoty. Za odlehlou hodnotu byl považován reakční čas přesahující průměrný reakční čas dané osoby o 2,5 směrodatné odchylky. Mezi muži a ženami nebyl zjištěn signifikantní rozdíl v reakčních časech, další analýza byla proto prováděna pro obě pohlaví společně. Efekt toxoplasmosy na reakční čas byl testován jednocestnou analýzou kovariance (kovariáta věk). Nenakažené osoby reagovaly v porovnání s osobami nakaženými signifikantně rychleji v 2. i 3. minutě testu ( F(1,113)=7,6, p=0,0067), respektive F(1,113)=4,8, p=0,030). Také celkový průměrný reakční čas nenakažených osob byl oproti osobám nakaženým signifikantně nižší (F(1,113)= 5,37, p=0,022). Přestože nenakažené osoby reagovaly rychleji i v 1. minutě testu, rozdíl hodnot nebyl v tomto případě signifikantní (F(1,113)=2,0, p=0,158).Výsledky všech testů a průměrné reakční časy Toxoplasma negativních a Toxoplasma pozitivních osob jsou uvedeny v tabulce č. 27. a zobrazeny v grafu č. 7.
|
reakční čas (milisec.) |
||||||||||
|
1. minuta |
2. minuta |
3. minuta |
1.-3. minuta |
|||||||
|
Toxoplasma negativní |
průměr |
254,9 |
258,5 |
265,4 |
259,9 |
|||||
|
sm. odch. |
34,2 |
27,6 |
27,7 |
27,3 |
||||||
|
Toxoplasma pozitivní |
průměr |
264,3 |
275,1 |
278,1 |
272,7 |
|||||
|
sm. odch. |
30,1 |
33,4 |
29,7 |
28,4 |
||||||
|
F |
2,02 |
7,62 |
4,81 |
5,37 |
||||||
|
df |
113 |
113 |
113 |
113 |
||||||
|
p |
0,158 |
0,007 |
0,030 |
0,022 |
||||||
Tab. č. 27: Průměrné reakční časy (v milisec.) v jednotlivých minutách a celkový průměrný reakční čas. ANCOVA.
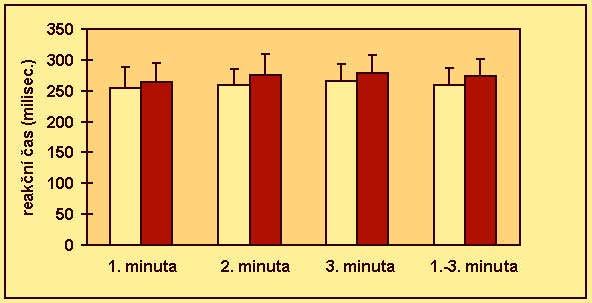
Graf č. 7: Průměrné reakční časy (v milisec.) v jednotlivých minutách a celkově. Chybové úsečky označují směrodatnou odchylku, světlé sloupce osoby Toxoplasma negativní a tmavé sloupce osoby Toxoplasma pozitivní.
Korelace mezi délkou infekce (odhadovanou podle hladin titrů protilátek) a průměrnými reakčnimi časy byla u 60 nakažených osob testována neparametrickým Kendallovým testem. Efekt věku byl eliminován použitím standartních residuálů z jednoduché regrese mezi věkem a průměrným reakčním časem. V první a druhé minutě testu byla zaznamenána významná negativní korelace mezi hladinou titrů protilátek a standartními residuály reakčních časů pro odpovídající věk (Kendall tau=-0,175, p=0,0384, resp.Kendall tau=-0,156, p=0,0477). Podobně i standartní residuály celkového průměrného reakčního času pro odpovídající věk signifikantně korelovaly s hladinou titrů protilátek (Kendall tau=-0.183, p=0,0285) (viz. graf č. 8). Přestože v třetí minutě testu byla korelace mezi standartními residuály reakčních časů pro odpovídající věk hodnocena jako nesignifikantní, byl zaznamenán stejný trend jako v ostatních minutách testu. Korelační koeficienty vypočtené Kendallovým testem a odpovídající hladiny signifikance jsou uvedeny v tab. 28.
|
N=60 |
Kendall tau |
p |
|
1. minuta |
-0,175 |
0,0384 |
|
2. minuta |
-0,156 |
0,0477 |
|
3. minuta |
-0,193 |
0,0767 |
|
1.-3. minuta |
-0,183 |
0,0285 |
Tab. č. 28: Korelační koeficient Kendall tau pro korelaci mezi hladinou titrů protilátek (reflektující délku infekce) a standartními residuály reakčních časů pro odpdovídající věk. Kendallův neparametrický test.
Graf č. 8: Korelace titrů protilátek (reflektující délku infekce) a standartních residuálů průměrných reakčních časů pro odpovídající věk v 1.-3. minutě testu.
5.5. Prevalence toxoplasmosy u účastníků dopravních nehod
Z celkového souboru 169 osob bylo z další analýzy vyloučeno 25 osob, kvůli vysokému věku (nad 70 let), 12 osob u nichž nebylo známo jakým způsobem se na dopravní nehodě podílely, 14 osob, které se na dopravní nehodě podílely jiným způsobem než jako chodci nebo řidiči (spolujezdci, cyklisté), 8 osob (řidičů), které nehodu nezavinily (byly nabouráni), 2 osoby u nichž byl zjištěn v krvi alkohol a 2 osoby-cizí státní příslušníci. Vlastní soubor tedy tvořilo 106 osob (47 chodců a 59 řidičů). Na základě výsledků serologického vyšetření na toxoplasmosu bylo 40 osob hodnoceno jako Toxoplasma pozitivní. Protože mezipohlavní rozdíly v prevalenci toxoplasmosy nebyly zjištěny, byla dále obě pohlaví hodnocena společně. Prevalence toxoplasmosy byla vyšší u řidičů ve srovnání s chodci, nicméně tento rozdíl nebyl významný a další analýza byla proto prováděna pro obě skupiny dohromady. Prevalence toxoplasmosy u účastníků dopravních nehod byla porovnána s prevalencí toxoplasmosy v normální populaci. Jako kontrolní soubor byly použity výsledky serologických vyšetření pro Prahu z let 1990 (Kodym a spol. 1995) a 1996 (Kodym a spol. 1997). Prevalence toxoplasmosy roste s věkem, byla proto provedena stratifikovaná analýza po jednotlivých věkových kategoriích. Celkově byl rozdíl v prevalenci toxoplasmosy mezi očekávanými hodnotami (serologické přehledy) a hodnotami skutečně zjištěnými (dopravní nehody) vysoce signifikantní (Mantel-Haenszel Chi-kvadrát=16,204, p=0,00006). Podle odhadu celkového váženého (maximum likelihood) odds ratio je mezi osobami Toxoplasma pozitivními 2,5 krát vyšší podíl účastníků dopravních nehod než mezi osobami Toxoplasma negativními. Pokud byly hodnoceny jednotlivé věkové kategorie zvlášť, byl pouze ve věkové kategorii 15-29 let podíl účastníků dopravních nehod mezi osobami nakaženými oproti nenakaženým významně (dolní mez konfidenčního intervalu vyšší než 1) vyšší. Četnosti nakažených a nenakažených osob v jednotlivých věkových kategoriích jsou uvedeny v tab. č. 29.
|
Toxoplasma negativní |
Toxoplasma pozitivní |
odds ratio |
konf. int. 95% |
|||||
|
věková kategorie |
dopravní nehody |
kontroly |
dopravní nehody |
kontroly |
||||
|
15-29 |
30 (60%) |
211 (84,7%) |
20 (40%) |
38 (15,3%) |
3,70 |
1,91-7,18 |
||
|
30-44 |
11 (73,3%) |
52 (77,6%) |
4 (26,7%) |
15 (22,4%) |
1,26 |
0,35-4,54 |
||
|
45-59 |
15 (57,7%) |
68 (75,6%) |
11 (42,3%) |
22 (24,4%) |
2,27 |
0,91-5,66 |
||
|
60-70 |
10 (66,7%) |
47 (77%) |
5 (33,3%) |
14 (23%) |
1,68 |
0,49-5,73 |
||
|
celkem |
66 (62,3%) |
378 (80,9%) |
40 (37,3%) |
89 (19,1%) |
2,5 |
1,63-4,06 |
||
Tab. č. 29: Četnosti nakažených a nenakažených osob v jednotlivých věkových kategoriích a celkově, v závorce uvedeny relativní četnosti. Pro každou věkovou kategorii je uvedeno odds ratio, celkové vážené (maximum likelihood) odds ratio a 95% konfidenční intervaly.
5.6. Vliv latentní toxoplasmosy na subjektivní hodnocení zdravotního stavu
Analýzou jednotlivých kontingenčních tabulek byl zjištěn signifikantní efekt toxoplasmosy na subjektivní hodnocení fyzického zdraví (Chi-kvadrát=3,882, df=1, p=0,049). Mezi osobami, které uváděly, že se obvykle necítí zdrávy, byla frekvence nakažených osob 41,7%, zatímco mezi zdravými osobami byla frekvence nakažených 23,6%. Četnosti a relativní četnosti odpovědí jsou uvedeny v tab. č. 30. Vzhledem k tomu, že prevalence toxoplasmosy stoupá s věkem, mohla by být zjištěná asociace mezi toxoplasmosou a hodnocením zdravotního stavu způsobena efektem věku. K odhalení vlastního efektu toxoplasmosy byla použita logistická regrese, kdy jako nezávislá binární proměnná vstupoval faktor toxoplasmosa a spojitá proměnná věk, jako závislá binární proměnná faktor zdravotní stav. Po přidání věku do analýzy jako další proměnné došlo k poklesu efektu toxoplasmosy pod hranici signifikance (t(322)=-1,824, p=0,007). Vzhledem k tomu, že byl testován efekt infekce (toxoplasmosy), bylo na místě očekávat spíše negativní hodnocení zdravotního stavu a bylo vhodnější použít spíše jednostranný test namísto dvoustranného. Při použití jednostranného testu zůstávala výsledná asociace mezi toxoplasmosou a subjektivním hodnocením zdravotního stavu stále signifikantní (p=0,035). Výsledky logistické regrese jsou uvedeny v tab. č. 31.
|
dobrý zdravotní stav |
||
|
ne |
ano |
|
|
Toxoplasma negativní |
14 (58,3%) |
230 (76,4%) |
|
Toxoplasma pozitivní |
10 (41,7%) |
71 (23,6%) |
Tab. č. 30: Frekvenční tabulka pro faktory zdravotní stav a toxoplasmosa. V závorce uvedeny relativní četnosti ve sloupcích.
|
konst.B0 |
věk |
toxoplasmosa |
|
|
odhad b |
3,242 |
-0,018 |
-0,799 |
|
SE |
0,5444 |
,0181 |
0,438 |
|
t(332) |
5,956 |
-0,98 |
-1,824 |
|
p |
0,000 |
0,328 |
0,035 |
Tab. č. 31: Asociace toxoplasmosy s faktorem zdravotní stav a matoucí proměnnou věk. V druhém sloupci je uveden odhad konstanty B0. Uveden je standartizovaný regresní koeficient b, střední chyba koeficientu b, hodnota t testu a odpovídající hladina signifikance, pro faktor zdravotní stav jednostranný test. Model logistická regrese.
Asociace mezi toxoplasmosou a psychickou pohodou nebyla zaznamenána (Chi-kvadrát=0,492, df=1, p=0,483). Ve skupině osob uvádějících, že se obvykle necítí psychicky dobře, byla frekvence Toxoplasma pozitivních osob 20,5%, na rozdíl od skupiny osob cítících se obvykle v psychické pohodě, mezi nimiž byla frekvence Toxoplasma pozitivních osob 25,4%. Četnosti a relativní četnosti odpovědí jsou uvedeny v tab. č. 32.
|
psychická pohoda |
||
|
ne |
ano |
|
|
Toxoplasma negativní |
35 (79,5%) |
203 (74,6%) |
|
Toxoplasma pozitivní |
9 (20,5%) |
69 (25,4%) |
Tab. č. 32: Frekvenční tabulka pro faktory psychická pohoda a toxoplasmosa. V závorce uvedeny relativní četnosti ve sloupcích.
U otázky na častost onemocnění byla z důvodu malého počtu odpovědí v některých kategoriích zredukována původní pěti stupňová škála na škálu dvoustupňovou. Vzniklá redukovaná proměnná měla tyto dvě hladiny: vůbec ne - jednou za dva roky a jednou za rok - častěji. Mezi skupinou nakažených a nenakažených osob nebyl zjištěn rozdíl v častosti onemocnění (hodnoceno v redukované formě) (Chi-kvadrát=0,199, df=1, p=0,656). Frekvence Toxoplasma pozitivních osob byla ve skupině osob uvádějících, že onemocní běžnými chorobami (chřipka, angína) nejvýše jednou za dva roky 26,3%, zatímco frekvence Toxoplasma pozitivních osob byla ve skupině uvádějící, že onemocní jednou za rok a častěji 24,1%. Četnosti a relativní četnosti odpovědí jsou uvedeny v tab. č. 33.
|
A |
častost onemocnění |
||||
|
vůbec ne |
za pět let |
za dva roky |
za rok |
častěji |
|
|
Toxoplasma negativní |
7 (87,5%) |
22 (61,1%) |
55 (78,6%) |
107 (77,0%) |
60 (74,1%) |
|
Toxoplasma pozitivní |
1 (12,5%) |
14 (38,9%) |
15 (21,4%) |
32 (23,0%) |
21 (25,9%) |
|
B |
častost onemocnění (reduk.) |
|
|
vůbec ne-za dva roky |
za rok-častěji |
|
|
Toxoplasma negativní |
84 (73,7%) |
167 (75,9%) |
|
Toxoplasma pozitivní |
30 (26,3%) |
53 (24,1%) |
Tab. č. 33: Frekvenční tabulka pro faktory častost onemocnění a toxoplasmosa. V závorce uvedeny relativní četnosti ve sloupcích. Tab. 33A původní forma a tab. 33B redukovaná forma.
Mezi toxoplasmosou a alergickými onemocněními byla zjištěna asociace blížící se hranici signifikance (Chi-kvadrát=3,783, df=1, p=0,052). Mezi osobami udávajícími, že netrpí alergickými chorobami byla frekvence Toxoplasma pozitiních osob 27,7%, zatímco mezi osobami, které uváděly, že trpí alergiemi, byla frekvence Toxoplasma pozitiních osob pouze 17,4%. Četnosti a relativní četnosti odpovědí jsou uvedeny v tab. č. 34.
|
alergie |
|||
|
ne |
ano |
||
|
Toxoplasma negativní |
175 (72,3%) |
76 (82,6%) |
|
|
Toxoplasma pozitivní |
67 (27,7%) |
16 (17,4%) |
|
Tab. č. 34: Frekvenční tabulka pro faktory alergie a toxoplasmosa. V závorce uvedeny relativní četnosti ve sloupcích.
5.7. Vliv latentní toxoplasmosy na index tělesné hmotnosti
Na základě výsledků serologického vyšetření metodou KFR bylo 31 mužů a 28 žen hodnoceno jako Toxoplasma pozitivní a 37 mužů a 19 žen jako Toxoplasma negativní. Průměrná tělesná výška byla u Toxoplasma negativních mužů 178,9 cm. a 167,5 cm. u žen. U Toxoplasma pozitivních mužů byla zjištěna průměrná tělesná výška 179,5 cm. a u žen 167,5 cm. Průměrná hmotnost u Toxoplasma negativních mužů byla 84,1 kg. a 64,9 kg u žen. U Toxoplasma pozitivních mužů byla zjištěna průměrná hmotnost 84,1 kg. a u Toxoplasma pozitivních žen 69,8 kg. Existence rozdílů v indexu tělesné hmotnosti (BMI) mezi nakaženými a nenakaženými osobami byla testována jednocestnou analýzou kovariance (kovariáta věk) pro obě pohlaví odděleně. Průměrný BMI u nenakažených mužů byl 26,2 zatímco u nakažených 26,1, což je rozdíl zcela nevýznamný. Také u žen nebyl zjištěn významný rozdíl v BMI mezi nakaženými a nenakaženými osobami. Průměrný BMI u nenakažených žen činil 23,3 zatímco u nakažených žen 24,9. Výsledky testů a všechny naměřené hodnoty jsou uvedeny v tab. č. 35. pro muže a v tab. č. 36 pro ženy.
V další části analýzy bylo zjišťována existence korelace mezi délkou infekce, odhadovanou na základě hladin titrů protilátek, a BMI. Efekt věku byl eliminován použitím standartních residuálů z jednoduché lineární regrese mezi věkem a BMI. Kendallovým neparametrickým testem nebyla zjištěna korelace mezi hladinou titrů protilátek a BMI a to jak u mužů tak i u žen. Korelační koeficienty vypočtené Kendallovým testem a odpovídající hladiny signifikance jsou uvedeny v tab. č. 37.
|
muži |
výška (cm) |
hmotnost (kg) |
BMI |
|
|
Toxoplasma negativní |
průměr |
178,9 |
84,1 |
26,2 |
|
sm. odch. |
6,7 |
14,5 |
3,9 |
|
|
Toxoplasma pozitivní |
průměr |
179,5 |
84,1 |
26,1 |
|
sm. odch. |
6,1 |
13,4 |
3,8 |
|
|
F |
0,179 |
|||
|
p |
0,673 |
Tab. č. 35: Tělesná výška, hmotnost a index tělesné hmotnosti (BMI) u mužů. ANCOVA.
|
ženy |
výška (cm) |
hmotnost (kg) |
BMI |
|
|
Toxoplasma negativní |
průměr |
167,5 |
64,9 |
23,3 |
|
sm. odch. |
6,8 |
8,5 |
3,7 |
|
|
Toxoplasma pozitivní |
průměr |
167,5 |
69,8 |
24,9 |
|
sm. odch. |
6,9 |
11,9 |
4,2 |
|
|
F |
0,652 |
|||
|
p |
0,424 |
Tab. č. 36: Tělesná výška, hmotnost a index tělesné hmotnosti (BMI) u žen. ANCOVA
|
N |
Kendall tau |
p |
|
|
muži |
31 |
-0,166 |
0,188 |
|
ženy |
28 |
-0,003 |
0,982 |
Tab. č. 37: Korelační koeficient Kendall tau pro korelaci mezi hladinou titrů protilátek (reflektující délku infekce) a standartními residuály BMI pro odpdovídající věk. Kendallův neparametrický test.
6. Diskuse
6.1. Analýza epidemiologických faktorů zvyšujících riziko infekce Toxoplasma gondii
Analýzou jednotlivých epidemiologických faktorů byla zjištěna asociace mezi toxoplasmosou a chovem kočky, konzumací syrového masa, chovem králíků a velikostí sídla v němž tázaný subjekt prožil mládí. Výsledky analýzy interakcí však ukazují, že asociace mezi velikostí sídla a toxoplasmosou zjištěná jednoduchou analýzou kontingenčních tabulek je spíše důsledkem interakce mezi faktory velikost sídla a konzumace syrového masa.
Velmi významnou roli z hlediska infekce toxoplasmou hraje v české populaci konzumace syrového masa. Toto zjištění je ve shodě s jinými českými (Flegr, a spol. 1998) i zahraničními studiemi (Braveny a spol. 1973; Stagno & Thiermann 1973; Buffolano a spol. 1996). Ve studiích z Panamy (Frenkel a spol.1995),La Guadaloupe (Barbier a spol. 1983) a Švédska (Huldt a spol. 1979) však asociace mezi konzumací syrového masa a toxoplasmosou nebyla zjištěna.. To může být odrazem odlišných potravních zvyklostí v populacích, v nichž byly tyto studie prováděny. Pokud by v těchto zemích byl zvyk konzumovat pouze syrové hovězí maso, bylo by riziko infekce, vzhledem k nízké prevalenci toxoplasmosy u skotu (Hejlíček & Literák 1992), velmi malé. Podobná vysvětlení však tyto studie neuvádějí.
Podle výsledků této práce zvyšuje konzumace syrového masa riziko infekce 19 krát. Zjištění takto vysokého rizika může být částečně vysvětleno tím, že v otázce bylo zahrnuto nejen jezení syrového masa, ale i ochutnávání při vaření. Dalo by se předpokládat, že zvyk ochutnávat syrové maso při vaření se bude častěji vyskytovat u žen. Mezipohlavní rozdíly však zjištěny nebyly. Významná interakce mezi konzumací syrového masa a velikostí sídla ukazuje, že konzumace syrového masa se uplatňuje jako rizikový faktor především na venkově. Tento jev by mohl být důsledkem rozdílů v potravních zvyklostech mezi venkovem a velkými městy. Ve velkých městech se syrové maso jí často ve formě tatarských bifteků vyráběných z mraženého masa. Mražení je pro infekční agens-tkáòové cysty letální (Jíra & Rosický 1983), takže riziko nákazy při takovýchto praktikách je minimální. Na druhou stranu na venkově tvoří značný podíl jídelníčku maso z vlastních chovů. Je známo, že promořenost jatečných zvířat je v České republice nejvyšší právě v domácích chovech (např. 35,1% u prasat) (Hejlíček & Literák 1993b). Ochutnávání syrového masa z vlastního chovu při vaření nebo konzumace špatně tepelně upravených domácích produktů (např. klobás) je pak významným zdrojem infekce.
Riziko infekce také významně zvyšuje chov kočky. Na druhou stranu asocice mezi frekvencí kontaktu s kočkou a toxoplasmosou zjištěn nebyl. Výsledky ostatních studií se v tomto ohledu různí. V studiích z Ruska (Ševkunova a spol. 1976) a La Guadeloupe (Barbier a spol. 1983) byla zjištěna vyšší frekvence toxoplasmosy mezi chovateli koček. Naproti tomu v studiích z Itálie (Buffolano a spol. 1996) a Švédska (Huldt a spol. 1979) asociace mezi chovem kočky a toxoplasmosou nebyla zjištěna. Podobně rozporné jsou výsledky studií, které se probandů dotazovaly na kontakt s kočkou. Frenkel a Ruiz (1980) zjistili v Kostarice vyšší prevalenci toxoplasmosy u osob, které udávaly kontakt s kočkou. Naproti tomu studie z Německa (Braveny a spol. 1973) podobnou asociaci nezjistila. Také v Japonsku nebyla asociace mezi frekvencí kontaktu s kočkou a toxoplasmosou zjištěna (Konishi & Takahashi 1987). Většina studií však uvažovala pouze jeden typ expozice. Pouze v jedné studii bylo sledováno více typů expozice, nicméně v žádném ze sledovaných typů nebyl vztah s toxoplasmosou zaznamenán (DiGiacomo a spol. 1990).
Zjištěná asociace mezi toxoplasmosou a chovem kočky, nikoliv však frekvencí kontaktu s kočkou ukazuje na nutnost zohledňování různých typů expozice rizikovému faktoru. Dobré srovnání poskytuje česká studie (Flegr a spol. 1998) založená na obdobném populačním vzorku jako tato práce. Autoři se probandů v jediné otázce tázali, zda často hladí kočku nebo zda chovají (případně chovali) kočku. Na základě odpovědí na jedinou takto položenou otázku došli autoři k závěru, že kontakt s kočkou nezvyšuje pravděpodobnost infekce toxoplasmou. Je tedy vidět, že způsob, jakým jsou otázky v epidemiologických výzkumech formulovány, může zásadním způsobem ovlivnit výsledky těchto studií. Vzájené srovnání s ostatními pracemi ztěžuje fakt, že většina studií neuvádí otázky, jež byly probandům předkládány (Flegr a spol. 1998).
K infekci chovatelů koček by mohlo docházet při manipulaci s nádobou, do níž kočky defekují. Zdrojem infekce chovatele by však také mohlo být syrové maso připravované jako krmivo pro kočky. Je známo, že parazit může pronikat do hostitele v místech drobných poranění (Jíra & Rosický 1983). Na druhou stranu pokud je kočka držena pouze v bytě, defekační nádoba je denně vyměňována (oocysty začnou být infekční teprve po třech dnech) a je krmena pouze konzervovaným masem, neměla by představovat vážný zdroj infekce (Frenkel, Dubey 1972).
Pravděpodobnost infekce Toxoplasma gondii je také zvýšena u chovatelů králíků. Vzhledem k tomu, že promořenost králíků v malochovech je velmi vysoká (53%) (Hejlíček & Literák 1994), není toto zjištění příliš překvapující. Vyšší frekvence toxoplasmosy u chovatelů králíků byla zjištěna i v jiných studiích (Zástěra a spol. 1966). Chovatelé se zřejmě nakazí oděrkami v kůži při zpracování králičího masa nebo při konzumaci špatně tepelně upraveného masa. Nicméně asociace mezi chovem králíků a konzumací syrového masa zjištěna nebyla.
Chov psa zřejmě nehraje v české populaci z hlediska epidemiologie toxoplasmosy významnou roli. K podobným závěrům došly i jiní autoři (Ševkunova a spol. 1975). Na druhou stranu v studii z Panamy bylo zjištěno, že chov psa zvyšuje riziko infekce (Frenkel a spol. 1995). Autoři se domnívají, že pes hraje významnou roli jako mechanický přenašeč, neboť je u něj častá koprofílie.
Zajímavým zjištěním je významná interakce mezi faktory chov kočky a konzumace syrového masa. U této interakce hodnota odds ratio nižší než jedna naznačuje, že by v souboru mohla existovat skupina osob, které záměrně či nezáměrně na obě otázky nepravdivě odpovídají kladně. Existence takové skupiny osob byla zjištěna i v předešlé studii (Flegr a spol. 1998).
6.2. Analýza asociací faktorů osobnosti s epidemiologickými faktory zvyšujícími riziko infekce Toxoplasma gondii
Zjištěné asociace mezi některými faktory osobnosti a epidemiologickými faktory, jež zvyšují riziko nákazy, naznačují, že osobnost jedince může ovlivnit pravděpodobnost nákazy toxoplasmou. Osobnostní faktory, které jsou asociovány s rizikovými faktory, jsou však různé u mužů a u žen. Toto zjištění není překvapivé, protože mezipohlavní rozdíly byly zjištěny prakticky u všech osobnostních faktorů monitorovaných Cattellovým dotazníkem (Havlíček & Flegr nepublikovaná data). Ukazuje se, že pro muže chovatele, a to jak koček, tak i králíků, je typická především závislost na skupině (nízké skóre ve faktoru Q2). Byly zjištěny i další asociace mezi epidemiologickými faktory a faktory osobnosti. Nicméně pouze rozdíly ve fraktoru Q2 mezi muži chovateli a nechovateli králíků zůstávaly signifikantní po Bonfferroniho korekci na mnohonásobné porovnávání. Použití korekce bylo v tomto případě vhodné, neboť pro každý epidemiologický faktor bylo provedeno 16 testů (testováno 16 faktorů osobnosti) a získání signifikantního výsledku jako důsledek náhody tak značně překračoval hladinu 5%.
Přestože rizikové faktory, zejména chov kočky a králíků jsou spojeny s určitými faktory osobnosti, jedná se o jiné osobnostní faktory než ty, ve kterých byly zjištěny rozdíly mezi nakaženými a nenakaženými osobami. Pouze v případě faktoru A byla v souboru žen zjištěna asociace tohoto faktoru s rizikovým faktorem chov králíků blížící se hranici signifikance (p=0,067). Pokud však vezmeme v úvahu počet provedených testů (16) je zřejmé, že jde nejspíše o náhodný efekt. Na základě těchto výsledků je tedy možné vyloučit, že rozdíly v psychologickém profilu osob Toxoplasma pozitivních a Toxoplasma negativních jsou příčinou infekce.
6.3. Změny osobnosti v důsledku latentní toxoplasmosy
Na základě srovnání osobnostních profilů nakažených a nenakažených dárců krve je možno potvrdit existenci rozdílů v profilu osobnosti Toxoplasma negativních a Toxoplasma pozitivních osob, zjištěných v předešlých studií (Flegr & Hrdý 1994; Flegr a spol. 1996; Flegr & Havlíček 1999). V této práci byla zaměřena pozornost především na ty faktory osobnosti u nichž je známo, že vykazují rozdíly mezi nakaženými a nenakaženými osobami. Protože byl znám směr změn v těchto faktorech, byl použit jednostranný test namísto dvoustranného. Rozdíl mezi nakaženými a nenakaženými osobami byl zjištěn ve faktorech G (síla superega) a N (naivita) u mužů a ve faktorech A (afektothýmie) a N u žen. Právě faktor G u mužů a faktor A u žen vykazovaly největší rozdíly v souboru biologů (Flegr a spol. 1996). Navíc u těchto faktorů je známo, že míra jejich změny koreluje s délkou infekce (Flegr a spol. 1996; Flegr a spol. zasláno do tisku). Nicméně po provedení Bonfferroniho korekce pro mnohonásobná porovnání zůstával významný pouze efekt ve faktoru A u žen. Jak již bylo uvedeno výše, při provádění většího množství testů je nutné počítat s tím, že roste pravděpodobnost chyby 1. druhu (Lepš 1996). Na druhou stranu Bonfferroniho korekce je metoda velmi konzervativní a snižuje tak pravděpodobnost zamítnutí nulové hypotézy a tím pádem zvyšuje pravděpodobnost chyby 2. druhu. Zamítnutí nulové hypotézy, přestože platí (chyba 1. druhu), je však větší chybou, než její nezamítnutí, přestože neplatí (chyba 2. druhu) (Lepš 1996).
Rozdíly v osobnostním profilu Toxoplasma negativních a Toxoplasma pozitivních osob byly interpretovány jako výsledek působení parazita (Flegr a spol. 1996). Tento způsob interpretace podporuje jednak zjištění, že skóre v některých faktorech osobnosti, v nichž se liší nakažené osoby od nenakažených, koreluje s délkou infekce (Flegr a spol. 1996; Flegr a spol. zasláno do tisku). Zároveň nebyla zjištěna korelace mezi faktory zvyšujícími riziko infekce a faktory osobnosti v nichž se liší osoby Toxoplasma negativní a Toxoplasma pozitivní (viz výše).
Behaviorální změny způsobené toxoplasmou jsou známy i u hlodavců. Nakažené myši jsou hyperaktivní (Hay a spol. 1983b, Hay a spol. 1985), hůře rozpoznávají nové podněty od známých (Hutchison a spol. 1980b) a v sociálních vztazích jsou více agresivní (Arnott a spol. 1990) ve srovnání s kontrolami. U potkanů je infekce Toxoplasma gondii spojena se snížením neofobie (Webster a spol. 1994) a stejně jako u myší s hyperaktivitou (Webster 1994). Autoři všech uvedených prací se domnívají, že změny v chování hlodavců zvyšují pravděpodobnost ulovení nakažených zvířat kočkou a tím i přenosu toxoplasmy do konečného hostitele.
U současného člověka však změny osobnosti pravděpodobnost transmise nezvyšují (lze si jen těžko předtavit, že člověk se nechá sežrat kočkou). Je však známo, že predátorem evolučních předků člověka byly velké kočkovité šelmy (Beneš 1994). Protože konečným hostitelem toxoplasmy mohou být kromě kočky domácí i ostatní kočkovité šelmy čeledi Felidae (Jíra & Rosický 1983), nelze tedy zcela vyloučit, že v minulosti vedla manipulační aktivita toxoplasmy k zvýšení pravděpodobnosti přenosu. Biologické interpretace změn v osobnosti ztěžuje však fakt, že v mnoha osobnostních faktorech mají tyto změny opačný směr u mužů a u žen.
V souboru biologů nebyly zaznamenány po pěti letech systematické změny ve faktorech osobnosti, ve kterých byly pozorovány rozdíly mezi nakaženými a nenakaženými osobami při původním testování. Nicméně výsledky získané v tomto souboru osob je obtížné interpretovat, protože naprostá většina osob věděla při opakovaném testování, zda jsou či nejsou nakaženi. Tato znalost mohla zásadně ovlivnit způsob vyplňování psychologického dotazníku. Osoby, u nichž byla tímto způsobem narušena naivita, se tak mohly stylizovat způsobem, jímž se domnívaly, že se od nich očekává. Navíc v souboru mužů odpovídali na dotazník častěji nakažení muži, což též mohlo určitým způsobem zkreslit výsledky. Dalším faktorem, který mohl ovlivnit výsledky, je případná změna „toxoplasma statusu“ některých osob. Opětné přetestování na toxoplasmosu nebylo z finančních a technických důvodů prováděno. Značná část osob již po pěti letech od původního testování nestudovala na univerzitě a byla tak osobně prakticky nedosažitelná. Protože parazit perzistuje doživotně v hostiteli (Dubey 1986), připadala v úvahu pouze infekce nenakažených osob. Jeden takový případ byl zaznamenán i v našem souboru (jednalo se o akutní toxoplasmosu). K výraznějšímu nárůstu prevalence toxoplasmosy po 20 roce života dochází pouze u mužů (o 8,3%). Většina žen se nakazí již v mladších věku (Kodym a spol. 1995). K infekci parazitem zřejmě došlo pouze u malého počtu osob (především mužů) z celého souboru. Ve srovnání s narušením naivity (viz výše) hrál nejspíše tento faktor pouze podružnou roli.
6.4. Vliv latentní toxoplasmosy na psychomotorický výkon
Výsledky psychomotorického testování dárců krve ukazují, že latentní toxoplasmosa je spojena se zhoršením psychomotorické výkonnosti. Nakažené osoby reagovaly na stimul signifikantně pomaleji v 2.a 3. minutě testu i v celkovém hodnocení. V tomto případě byl z důvodu určité opatrnosti použit dvoustranný test. Protože však byl testován efekt infekce, u níž se dá předpokládat spíše zhoršení výkonnosti, bylo by patrně vhodnější použít jednostranný test. Dosažená hladina významnosti by tak byla ještě dvakrát vyšší. Výkon v testu se zhoršoval s délkou infekce, odhadované na základě hladin titrů protilátek. Tento efekt byl zjištěn i přesto, že hladiny titrů protilátek jsou jen hrubým odhadem délky infekce (Holliman 1997). Použitý test je určen k zjišťování oko-motorické koordinace a udržení pozornosti. Nesignifikantní výsledek v 1. minutě testu naznačuje, že by v důsledku infekce mohlo docházet spíše k zhoršování pozornosti než oko-motorické koordinace.
V souboru biologů však významný efekt toxoplasmosy na psychomotorický výkon zjištěn nebyl. Tato diskrepance byla zřejmě způsobena narušením naivity ohledně „toxoplasma statusu“ v souboru biologů. Osoby v tomto souboru byly testovány na toxoplasmosu v rámci předešlých studií (Flegr & Hrdý 1994; Flegr a spol. 1996). Většina z nich tedy věděla, narozdíl od dárců krve, zda jsou nakažení či nenakažení. Tento faktor mohl závažným způsobem ovlivnit výsledky. Je proto nutné velké opatrnosti při interpretaci výsledků tohoto souboru. Nicméně i v tomto souboru byl zachován stejný trend čili osoby Toxoplasma pozitivní dosahovaly vyšších reakčních časů v porovnání s osobami Toxoplasma negativními.
Zhoršení psychomotorické výkonnosti je známo také u některých bakteriálních a virových nákaz. Experimentálně navozená infekce bakterií Pasteurella tularensis způsobovala celkové zhoršení psychomotorického výkonu o 25% (Alluisi a spol. 1971). Selektivní efekt na výkonnost byl zjištěn u infekcí viry způsobujícími onemocnění horních cest dýchacích, chřipku a infekční mononukleózu. Zatímco onemocnění horních cest dýchacích způsobuje zhoršení oko-motorické koordinace a udržování pozornosti, onemocnění chřipkou způsobuje zpomalení reakcí na stimulus objevující se v nepravidelných intervalech nebo na různých místech (Smith a spol. 1989; Hall & Smith 1996b). Akutní fáze onemocnění infekční mononukleózou je spojena se zhoršením reakčního času a udržení paměti, oproti tomu chronická fáze infekce je spojena se zhoršením krátkodobé paměti a logického zdůvodňování (Hall & Smith 1996a).
V této práci byly použity dva testy. V souboru dárců krve byl, z důvodu minimalizace zatížení provozu transfusní stanice, použit pouze jeden test. Oba testy však zjišťují zároveň dvě různé funkce: oko-motorickou koordinaci a udržení pozornosti. Nelze tedy na základě těchto výsledků říci, zda i latentní toxoplasmosa má spíše selektivní efekt na psychomotorickou a mentální výkonnost, nebo zda dochází k celkovému zhoršení psychických funkcí. K rozhodnutí této otázky by bylo nutné použít celou baterii testů měřících jednotlivé psychomotorické a kognitivní funkce.
Mechanismy, na jejichž základě dochází v důsledku infekce k zhoršení výkonnosti, nejsou známy. Latentní toxoplasmosa má ve valné většině případů minimální, případně žádné klinické následky. Nelze tedy předpokládat, že zjištěné změny jsou produktem onemocnění. Přestože parazit perzistuje v hostiteli doživotně, většina lidí neví o tom, že je nakažena. U myší infikovaných toxoplasmou byly zjištěny změny v koncentracích některých neurotransmiterů (Stibbs 1995). Je tedy možné, že parazit určitým způsobem zasahuje do produkce některých neuroaktivních látek. Takový zásah by mohl vést i k pozorovaným behaviorálním změnám.
6.5. Prevalence toxoplasmosy u účastníků dopravních nehod
Zjištěná vyšší prevalence toxoplasmosy u účastníků dopravních nehod ve srovnání s normální populací naznačuje, že infekce parazitem Toxoplasma gondii hraje v těchto situacích významnou roli. Po odstranění efektu rozdílné věkové skladby obou porovnávaných souborů bylo zjištěno, že mezi nakaženými osobami byl 2,5 krát vyšší podíl účastníků dopravních nehod než mezi osobami nenakaženými.
Příčinou dopravní nehody může být celá řada faktorů. Naší snahou bylo zařadit do souboru pouze ty řidiče, kteří se na dopravní nehodě aktivně podíleli. Z tohoto důvodu byli z analýzy vyřazeni řidiči, kteří byli nabouráni, dále též osoby, které se dopravní nehody účastnily jako spolujezdci a nebo nebylo možné podle dostupných lékařských záznamů zjistit, jakým způsobem se na nehodě podílely. Vyřazeny byly též osoby, u nichž byl zjištěn alkohol v krvi a nebo nepocházely z české populace.
Mezi významné funkce uplatňujícími se v dopravních situaciích patří oko-motorická koordinace (např. při brždění, vyhybání se překážce a pod). A dále také pozornost věnovaná řízení, případně přecházení vozovky u chodců. V laboratorních testech bylo zjištěno (viz výše), že v důsledku latentní toxoplasmosy dochází k zhoršení těchto funkcí. Je tedy možné, že zjištěná vyšší prevalence toxoplasmosy u účastníků dopravních nehod ve srovnání s normální populací, je odrazem zhoršení psychomotorických funkcí v důsledku infekce.
Naše výsledky ukazují, že toxoplasmosa hraje podobnou roli při dopravních nehodách u řidičů i u chodců. Prevalence toxoplasmosy byla nesignifikantně nižší u chodců ve srovnání s řidiči. Tento jev by se dal vysvětlit tím, že chodci jsou ve srovnání s řidiči častěji pasivní obětí dopravní nehody než jejím viníkem a tím pádem se u chodců méně uplatňují psychomotorické funkce, které jsou parazitem ovlivněny. Nejvyšší podíl účastníků dopravních nehod mezi Toxoplasma pozitivními osobami byl podle odhadu odds ratio zjištěn v nejmladší sledované věkové kategorii (15-29 let). Přestože podobný trend byl zjištěn i v ostatních sledovaných věkových kategoriích, nebyl v těchto věkových kategoriích významný (dolní mez 95% konf. intervalu nižší než 1). Tento jev by mohl být důsledkem poměrně malého počtu osob ve vyšších věkových kategoriích. Je však také možné, že role toxoplasmosy v dopravních nehodách relativně klesá s věkem v souvislosti s růstem významu faktorů spojených s vlastním stárnutím. Faktory spojené se stárnutím (např. zhoršení zraku, motoriky, koordinace) jsou zřejmě primární příčinou nehod v nejvyšších věkových kategoriích. Z tohoto důvodu jsme již před zahájením pokusu rozhodli do něj nezařazovat osoby nad 70 let.
6.6. Vliv latentní toxoplasmosy na subjektivní hodnocení zdravotního stavu
Výsledky této části práce ukazují, že osoby Toxoplasma pozitivní hodnotí svůj zdravotní stav častěji negativně ve srovnání s osobami Toxoplasma negativními. Pokud byl do analýzy zavzat věk jako další proměnná došlo k poklesu významnosti efektu toxoplasmosy, nicméně při použití jednostranného testu zůstával stále signifikantní na hladině 5%. Jednostrannému testu byla v tomto případě dána přednost, protože byla testováno, zda toxoplasmosa je spojena s negativním hodnocením zdravotního stavu. Je nepravděpodobné, že by v důsledku onemocnění, byť latentního, docházelo k zlepšení zdravotního stavu. Zjištěná asociace mezi toxoplasmosou a subjektivním hodnocením zdravotního stavu nabízí možné alternativní vysvětlení rozdílů v psychologickém profilu mezi nakaženými a nenakaženými osobami. Parazit je schopen v hostiteli doživotně přežívat (Dubey 1986). Je tedy možné, že dlouhodobé negativní ovlivňování zdravotního stavu hostitele může vést až ke změnám v jeho osobnosti. Zjištěné rozdíly v některých faktorech osobnosti mezi nakaženými a nenakaženými osobami by pak byly spíše než výsledkem manipulační aktivity ze strany parazita, důsledkem zhoršení zdravotního stavu parazitovaného jedince.
Zajímavým zjištěním je negativní asociace mezi toxoplasmosou a alergickými onemocněními, která se blížila pětiprocentní hladině signifikance. Tento výsledek naznačuje, že by osoby s alergiemi tj. projevy hyperimunity, mohly vykazovat zvýšenou rezistenci vůči nákaze parazitem Toxoplasma gondii. Některé recentní studie (Bell 1996; Allen & Maizels 1996) poukazují na souvislost mezi alergiemi a výrazným snížením výskytu (téměř eradikaci) helmintárních infekcí v zemích s vysokou úrovní zdravotnictví a hygieny. Podle představ některých autorů (Bell 1996; Allen & Maizels 1996) hráli helminti důleležitou roli v evoluci homeostáze imunitního systému. Pokud je člověk vystaven od raných ontogenetických období přítomnosti helmintárních infekcí dochází k chronické aktivaci složek (především T-helpers2 buněk) imunitního systému odpovědných za regulaci imunitní odpovědi. V připadě, že imunitní systém není takto stimulován, tak jak tomu je ve vyspělých zemích, může v některých případech dojít k poruchám reaktivity čili alergickým onemocněním. Tuto hypotézu podporuje i fakt, že v rozvojových zemích, kde je helmintárními infekcemi postižena převážná část populace, se alergická onemocnění vyskytují velmi zřídka (Bell 1996). Při reakcích na protozoární infekce se však podílejí jiné části imunitního systému (Remington & Krahenbuhl 1982) a podobná souvislost s alergickými onemocněními není známa.
Při interpretaci našich výsledků je však nutná určitá opatrnost. Protože osoby byly testovány na toxoplasmosu v rámci předešlých studií (Flegr & Hrdý 1994; Flegr a spol. 1996), většina z nich věděla zda jsou nakaženi či nikoliv. Tato znalost mohla ovlivnit způsob jakým na položené dotazy odpovídaly.
6.7. Vliv latentní toxoplasmosy na index tělesné hmotnosti
Na základě našich výsledků nemůžeme potvrdit zjištění Flegra a Hrdé (zasláno do tisku), že v důsledku latentní toxoplasmosy dochází k poklesu tělesné hmotnosti. Je však třeba zdůraznit, že náš soubor byl význnamně menší oproti souboru použitém v původní studii (810 osob). Navíc studie Flegra a Hrdé (zasláno do tisku) sledovala hmotnost žen v 16. týdnu gravidity. Je možné, že pozorované rozdíly v hmotnosti mohly být způsobeny nižšími váhovými přírůstky Toxoplasma pozitivních žen v prvním trimestru gravidity. V současné české populaci ovlivňují tělesnou hmotnost především sociální faktory jako jsou potravní návyky, množství pohybových aktivit atd (Hájková 1997). Pokud by dopad toxoplasmosy na tělesnou hmotnost nebyl příliš výrazný, je možné, že byl tento efekt překryt významnějšími sociálními faktory. K odhalení efektu toxoplasmosy (pokud skutečně existuje) by bylo nutné použít většího souboru osob.
Pokles tělesné hmotnosti v důsledku infekce je znám u mnoha parazitárních onemocnění. Většina intestinálních helmintů (např. Ascaris lumbricoides, Trichuris trichiura, Ancylostoma duodenale a další), ale i protozoí (např. Gardia intestinalis, Entamoeba hystolitica a další) způsobuje poruchy trávení nebo absorbce nutričních složek potravy (Solomons 1993). Nutriční ztráty způsobené parazitem se pak negativně projeví i na tělesné hmotnosti parazitovaného jedince. Jakým způsobem by mohlo docházet k poklesu tělesné hmotnosti v důsledku latentní toxoplasmosy není známo.
6. Závěrečné shrnutí
Ř Riziko infekce parazitem Toxoplasma gondii zvyšuje konzumace syrového masa, chov kočky a chov králíků. Konzumace syrového masa se jako rizikový faktor uplatňuje především na venkově, zřejmě díky vysoké promořenosti jatečných zvířat z domácích chovů.
Ř Osobnostní profil jedince může ovlivnit riziko infekce toxoplasmou. Osobnostní faktory odpovědné za rizikové chování se však liší od těch, které vykazují rozdíly mezi nakaženými a nenakaženými osobami. Je tedy možné vyloučit, že rozdíly v osobnostním profilu Toxoplasma negativních a Toxoplasma pozitivních osob jsou příčinou infekce.
Ř Existence rozdílů v psychologickém profilu nankažených a nenakažených osob byla ověřena na nezávislém souboru osob. Rozdíly vykazovaly stejné osobnostní faktory jako v předešlých studiích. Systematické změny v psychologickém profilu u osob retestovaných po pěti letech se nepodařilo zjistit.
Ř Latentní toxoplasmosa zhoršuje výkonnost v psychomotorickém testu reakčního času. Navíc výsledek testu negativně koreluje s délkou infekce.
Ř Prevalence toxoplasmosy u účastníků dopravních je významně vyšší ve srovnání s normální populací. Jedná se zřejmě o důsledek negativního působení parazita na psychomotorické funkce.
Ř Osoby Toxoplasma pozitivní hodnotí ve srovnání s osobami Toxoplasma negativními častěji svůj zdravotní stav jako negativní. U nakažených osob se méně často vyskytují alergická onemocnění.
Ř Na základě našich výsledků nemůžeme potvrdit zjištění z předešlé studie, že latentní toxoplasmosa negativně ovlivňuje tělesnou hmotnost infikované osoby.
8. Použitá literatura
Adams, E.J., Stephenson, L.S., Latham, M.C., Kinoti, S.N. (1994): Physical activity and growth of Kenyan school children with hookworm, Trichuris trichiura and Ascaris lumbricoides infections are improved after treatment with albendazole. J. Nutr. 124: 1199-1206.
Allen, J.E., Maizels, R.M. (1996): Immunology of human helminth infection. Int. Arch. Allergy Immunol. 109: 3-10.
Alluisi, E.A., Thurmond, J.B., Coates, G.D. (1971): Behavioral effects of infectious diseases: respiratory Pasteurella tularensis in man. Percept. Mot. Skills 32: 647-668.
anonymus, (1995): Electronic manual for StatisticaŇ/w. 5.0 program. StatSoft. inc.
Arnott, M.A., Cassella, J.P., Aitken, P.P., Hay, J. (1990): Social interaction of mice with congenital Toxoplasma infection. Ann. Trop. Med. Par. 84: 149-156.
Barbier, D., Ancelle, T., Martin-Bouyer, G. (1983): Seroepidemiological survey of toxoplasmosis in La Guadeloupe, French West Indies. Am. J. Trop. Med. Hyg. 32: 935-942.
Bell, R.G. (1996): IgE, allergies and helminh parasites: A new perspective on an old conundrum. Immunol. Cell Biol. 74: 337-345.
Beneš, J. (1994): Člověk. Mladá fronta, Praha.
Berdoy, M., Webster, J.P., Macdonald, D.W. (1995): Parasite-alterd behaviour: is the effect of Toxoplasma gondii on Rattus norvegicus specific? Parasitology. 111: 403-409.
Bozděch, V., Moronfoye, I.S., (1974): A skin-testing survey with toxoplasmin on positivity in the different age- and sex groups of the normal urban population of Kaduna (Nigerie). Folia Parasit. 21: 67-70.
Braveny, I., Disko, R., Janssen, H. (1973): Untersuchungen zur Epidemiologie der Toxoplasmose. Dtsch. med. Wschr. 98: 535-539.
Buffolano, W., Gilbert, R.E., Holland, F.J., Fratta, D., Palumbo, F., Ades, A.D. (1996): Risk factors for recent toxoplasma infection in pregnant women in Naples. Epidemiol. Infect. 116: 347-351.
Burkinshaw, J., Kirman, B.H., Sorsby, A. (1953): Toxoplasmosis in relation to mental deficiency. Brit. Med. J. 702: 1-6.
Cantella, R., Colichon, A., Lopez, L., Wu, C., Goldfarb, A., Cuadra, E., Latorre, C., Kanashiro, R., Delgado, M., Piscoya, Z. (1974): Toxoplasmosis in Peru. Trop. geogr. Med. 26: 204-209.
Cattell, R.B. (1970): Handbook for the Sixteen Personality Factors Questionnaire (16PF): Champaign: Institut for personality and ability testing.
Connolly, K.J., Kvalsvig, J.D. (1993) Infection, nutrition and cognitive performance in children. Parasitology 107: S187-S200.
Cook, I., Derrick, E.H. (1961): The incidence of toxoplasma antibodies in mental hospital patients. Austral. Ann. Med. 10: 137-141.
Černý, Z., Fusková, E., Jalinková, D., Jirsenská, D., Pechová, H. (1995): Klinické zkušenosti z epidemického výskytu toxoplasmózy na Jižní Moravě na přelomu let 1993-1994. Abstrakta celostátní konference infekcionistů Havířov 1995: 12-13.
Dawkins, R. (1998): Sobecký gen. Mladá Fronta, Praha.
De Roever-Bonnet, H. (1967): Toxoplasmosis in Surinam (Netherland Guyana): A Serological Survey. Trop. geogr. Med. 19: 221-228.
De Roever-Bonnet, H. (1972): Toxoplasmis in tropical Africa. Trop. geogr. Med. 24: 7-13.
Desmonts, G., Couvreur, J. (1974): Congenital toxoplasmosis. A prospective study of 378 pregnancies. N. Engl. J. Med. 290: 1110-1116.
Develoux, M., Candolfi, E., Hanga-Doumbo, S., Kien, T., (1988): La Toxoplasmose au Niger. Bull. Soc. Path. Ex. 81: 253-259.
DiGiacomo, R.F., Harris, N.V., Huber, N.L., Cooney, M.K. (1990): Animal Exposure and Antibodies to Toxoplasma gondii in a University Population. Am. J. Epidemiol. 131: 729-733.
Dubey, J.P. (1986): Toxoplasmosis. J. Am. Vet. Med. Assoc. 189: 166-170.
Dubey, J.P., Miller, N.L., Frenkel, J.K. (1970a): Charakterization of the new fecal form of Toxoplasma gondii. J. Parasit. 56: 447-456.
Dubey, J.P., Miller, N.L., Frenkel, J.K. (1970b): The Toxoplasma gondii oocyst from cat feces. J. Exp. Med. 132: 636-662.
Dubey, J.P., Frenkel, J.K. (1976): Feline toxoplasmosis from acutely infected and the development of Toxoplasma oocyst. J. Protozool. 23: 537-546.
Dubey, J.P., Lindsay, D.S., Speer, C.A. (1998): Structure of Toxoplasma gondii tachyzoites, bradyzoites, and sporozoites and biology and development of tissue cysts. Clinic. Microbiol. Rev. 11: 267-299.
Edwards, J.C. (1988): The effect of Trichinella spiralis infection on social interactions in mixed groups of infected and uninfected male mice. Anim. Behav. 36: 529-540.
Elias, M.I., (1966): Beitrage zur Epidemiologie der Toxoplasmose. Z. Tropenmed. Parasit. 17: 87-99.
Fayomi, B., Duong, T.H., Karagirwa, A., Marquet, N., Djivoh, C., Combescot, C. (1987): Etude Serologique de la Toxoplasmose et de L’Hydatidose chez les Eleveurs de Bovins et le Personnel des Abattoirs au Benin. Med. Trop. 47: 149-151.
Feldman, H.A. (1954): Laboratory methods in current use for the study of toxoplasmosis. Adv. Opthalmol. 3: 1-11.
Fetter, V. (1967): Antropologie. Academia, Praha.
Fiore, M., Moroni, R., Alleva, E., Aloe, L. (1996): Schistosoma mansoni: Influence of Infection on Mouse Behavior. Experim. Parasitol. 83: 46-54.
Flegr, J., Hrdý, I. (1994): Influence of chronic toxoplasmosis on some human personality factors. Folia Parasitol. 41: 122-126.
Flegr, J., Zitková, Š., Kodym, P., Frynta, D. (1996): Induction of changes in human behaviour by the parasitic protozoan Toxoplasma gondii. Parasitology. 113: 49-54.
Flegr, J., Hrdá, Š., Tachezy, J. (1998): The role of psychological factors in questionaire-based studies on routes of human toxoplasmosis transmission. Centr. eur. J. publ. Hlth. 6: 43-48.
Flegr, J., Havlíček, J. (1999): Changes in the personality profile of young women with latent toxoplasmosis. Folia Parasitol. 46: 22-28.
Flegr, J., Kodym, P., Tolarová, V. (zasláno do tisku): Correlation of latent Toxopalasma gondii infection with personality changes in women.
Flegr, J., Hrdá, Š. (zasláno do tisku): Influence of latent toxoplasmosis on human health.
Frenkel, J.K., Dubey, J.P. (1972): Toxoplasmosis and its prevention in cats and man. J. Infect. Dis. 126: 664-673.
Frenkel, J.K., Nelson, B.M., Arias-Stella, J. (1975a): Immunosupression and toxoplasmic encephalitis. Human Pathol. 6: 97-111.
Frenkel, J.K., Ruiz, A., Chinchilla, M. (1975b): Soil survival of Toxoplasma oocyst in Kansas and Costa Rica. Am. J. Trop. Med. Hyg. 24: 439-443.
Frenkel, J.K., Ruiz, A., (1980): Human toxoplasmosis and cat contact in Costa Rica. Am. J. Trop. Med. Hyg. 29: 1167-1180
Frenkel, J.K., Ruiz, A., (1981): Endemicity of toxoplasmosis in Costa Rica. Am. J. Epidemiol. 113: 254-269.
Frenkel, J.K., Hassanein, K.M., Hassanein, R.S., Brown ,E., Thulliez, P., Quintero-Nuinez, R. (1995): Transmission of Toxoplasma gondii in Panama City, Panama: A five-year prospective cohort study of children, cats, rodents, birds, and soil. Am. J. Trop. Med. Hyg. 53: 458-468.
Freytag, H.W., Haas, H. (1979): Zur Psychopathologie der erworbenen Toxoplasmose. Nervenarzt. 50: 128-131.
Gandahusada, S. (1978): Serological study for antibodies to Toxoplasma gondii in Jakarta, Indonesia. Southeast Asian J. Trop. Med. Pub. Hlth. 9: 308-311.
Garcia, G.D. (1979): Toxoplasmosis y enfermedades mentales. Rev. Cub. Med. Trop. 31: 127-131.
Garcia, G.D. (1983): Reactividad a la toxoplasmina aby positividad al Dye-test aby fijacion del complemento en personas aparentemente sanas de las provincias Ciudad de La Habana aby La Habana. Rev. Cub. Med. Trop. 35: 338-344.
Gibson, C.L. (1956): Distribution of toxoplasma antobodies in comparable urban and rural groups. Pub. Hlth. Rep. 71: 1119-1123.
Godwin, J.D., Remington, J.S. (1973): Prevalence of antibodies to toxoplasma in a group of Ghanians. Trop. geogr. Med. 25: 293-296.
Gross, U. (1996): Toxoplasma gondii Research in Europe. Parasitology Today. 12: 1-4.
Hadidjaja, P., Bonang, E., Suyardi, M.A., Abidin, A.N., Ismid, I.S., Margono, S.S. (1998): The effect of intervention methods on nutritional status and cognitive function of primary school children infected with Ascaris lumbricoides. Am. J. Trop. Med. Hyg. 59: 791-795.
Hagiwara, T., Katsube, Y., Muto, T., Imaizuni, K., Daigo, Y. (1981): Experimental feline toxoplasmosis. Jpn. J. Vet. Sci. 43: 329-336.
Hájková, R. (1997): Změny tělesné stavby obézních dětí vybraných věkových kategorií v průběhu lázeňské léčby. Diplomová práce, PřF UK Praha.
Hall, S.R., Smith, A.P. (1996a): Behavioural effect of infectious mononucleosis. Neuropsychobiology. 33: 202-209.
Hall, S., Smith, A. (1996b): Investigation of the effects and aftereffects of naturally occuring upper respiratory tract illnesses on mood and performance. Physiol. Behav. 59: 569-577.
Hay, J., Aitken, P.P., Hutchison, W.M., Graham, D.I. (1983a): The Effect of congenital and adult-acquired Toxoplasma infections on motor performance of mice. Ann. Trop. Med. Par. 77: 260-277.
Hay, J., Hutchison, W.M., Aitken, P.P., Graham, D.I. (1983b): The effect of congenital and adult-acquired Toxoplasma infections on activity and responsiveness to novel stimulation in mice. Ann. Trop. Med. Par. 77: 483-495.
Hay, J., Hutchison, W.M., Aitken, P.P. (1983c): The effect of Toxocara canis infection on the behaviour of mice. Ann. Trop. Med. Par. 77: 543-544.
Hay, J., Aitken, P.P., Hair, D.M., Hutchison, W.M., Graham, D.I. (1984): The effect of congenital Toxoplasma infection on mouse activity and relative preference for exposed area over a series of trials. Ann. Trop. Med. Par. 78: 611-618.
Hay, J., Aitken, P.P., Arnott, M.A. (1985): The influence of congenital Toxoplasma infection on the spontaneous running activity of mice. Z. Parasitenkd. 71: 459-462.
Hay, J., Arnott, M.A., Aitken, P.P., Kendall, A.T. (1986): Experimental toxocariasis and hyperactivity in mice. Z. Parasitenkd. 72: 115-120.
Hejlíček, K., Literák, I. (1992): Occurence of toxoplasmosis and its prevalence in cattle in the south Bohemian region. Acta Vet. Brno 61: 195-206.
Hejlíček, K., Literák, I. (1993a): Toxoplazmóza jatečných ovcí. Veterinářství 43: 414-417.
Hejlíček, K., Literák, I. (1993b): Prevalence of toxoplasmosis in pigs in the region of south Bohemia. Acta Vet. Brno 62: 159-166.
Hejlíček, K., Literák, I. (1994): Toxoplazmóza jatečných králíků. Veterinářství 44: 112-113.
Hejlíček, K., Literák, I., Chalupa,B. (1993): Výskyt protilátek proti Toxoplasma gondii u dárců krve za časové období 1980-1990. Čs. Epidem. 42: 135-140.
Holliman, R.E. (1997): Toxoplasmosis, behaviour and personality. J. Infect. 35: 105-110.
Holmes, J.C., Bethel, W.M. (1972): Modification of intermediete host behaviour by parasites. In: Canning, E.U., Wright, C.A (eds.) Behavioural aspects of parasite transmission. Suppl. 1 Zool. J. Linn. Soc. 52: 123-149.
Holmes, J.C., Zohar, S. (1990): Pathology and host behaviour. In: Barnard,C.J., BehnkeJ.M. (eds.) Parasitism and host behaviour. Taylor and Francis, London, New York, Philadelphia, 34-63.
Hoschl, C. Balon, R. (1980): Výsledky psychiatrického vyšetřování nemocných toxoplasmózou. Čas. Lék. čes. 119: 366-368.
Hosmer, D.W., Lemeshow, S. (1989): Applied logistic regression. Wiley, New York.
Huldt, G., Lagercrantz, R., Sheehe, P.R., (1979): On the epidemiology of human toxoplasmosis especially in children. Acta Paediatr. Scand. 68: 745-749.
Hutchison, W M. (1965): Experimental transmission of Toxplasma gondii. Nature 206: 961.
Hutchison, W.M., Work, K. (1969): Toxoplasma - A versatile parasite. New Scientist 46: 464-466.
Hutchison, W.M., Dunachie, J.F., Ferguson, D.J.P., Gardner, C.I. (1971): Endogenous development of the coccidan parasite Toxoplasma gondii. Trans. R. Soc. Trop. Med. Hyg. 65: 429-430.
Hutchison, W.M., Aitken, P.P., Wells, B.W.P. (1980a): Chronic Toxoplasma infections and familiarity-novelty discrimination in the mouse. Ann. Trop. Med. Par. 74: 145-150.
Hutchison, W.M., Bradley, M., Cheyne, W.M., Wells, B.W.P. Hay, J. (1980b): Behavioural abnormalities in Toxoplasma-infected mice. Ann. Trop. Med. Par. 74: 337-345.
Jankó, M., Czeizel, E. (1970): Epidemiology of toxoplasmosis in Hungary. Parasit. Hung. 3: 119-132.
Janků, J. (1923): Pathogenesa a patologická anatomie tak zvaného vrozeného kolombu žluté skvrny v oku normálně velikém a mikrophthalmickém s nálezem parazitů v sítnici. Čas. Lék. čes. 62: 1021-1027, 1054-1059, 1081-1085, 1111-1115, 1138-1143.
Jíra, J., Rosický B. (1983): Imunodiagnostika a epidemiologie toxoplasmosy. Academia, Praha.
Jírovec, O., Vojtěchovský, M. (1957): Zkušenosti s toxoplasminovým testem u psychiatrických pacientů. Hraniční problémy psychiatrie. 167: 67-76.
Jírovec, O. (1974): Lékařská parazitologie. Avicenum, Praha.
Johnson, A.M, Roberts, H., McDonald, P.J. (1980): Age-sex distribution of Toxoplasma antibody in the South Australian population. J. Hyg. Camb. 84: 315-320.
Kahn, H.A., Sempeos, Ch.T. (1989): Statistical methods in epidemiology. Oxford University Press, New York.
Kavaliers, M., Colwell, D.D. (1995a): Reduced spatial learning in mice infected with the nematode, Heligmosmoides polygyrus. Parasitology 110: 591-597.
Kavaliers, M., Colwell, D.D. (1995b): Discrimination by female mice between the odours of parasitized and non-parasitized males. Proc. R. Soc. Lond. B 261: 31-35.
Kleinbaum, D.G. (1994): Logistic regression. A self-learning text. Springer-Verlag.
Ko, R.C., Wong, F.W.T., Todd, D., Lam, K.C. (1980): Prevalence of Toxoplasma gondii antibodies in the Chinese population of Hong Kong. Trans. Royal Soc. Trop. Med. Hyg. 74: 351-354.
Kodym, P., Malý, M., Švandová, E., Zástěra, M. (1995): Incidence a séroprevalence toxoplasmosy v České republice v letech 1977-1994. Interní zprávy SZÚ, Praha.
Kodym, P., Malý, M., Švandová, E., Zástěra, M. (1997): Incidence a séroprevalence toxoplasmosy v České republice (Praha) v roce 1996. Interní zprávy SZÚ, Praha.
Kodym, P., Tolarová, V. (1998): Laboratorní diagnostika toxoplasmosy. Remedia - Klinická mikrobiologie. 7: 224-226.
Komiya, Y., Kobayashi, A., Koyama, T. (1961): Human toxoplasmosis, particularly on the possible source of its infection in Japan: a Rewiev. Jpn. J. Med. Sc. Biol. 14: 157-172.
Konishi, E., Takahashi, J. (1987): Some epidemiological aspects of Toxoplasma infection in a population of farmers in Japan. Inter. J. Epidemiol. 16: 277-281.
Koppe, J.G., Rothova, A. (1989): Congenital toxoplasmosis. Inter. Opthal. 13: 387-390.
Kouba, K., Štafová, J., Nevařilová, A., Flusser, J. (1966): Získaná toxoplasmóza probíhající pod obrazem psychiatrických poruch a myokarditidy. Čas. Lék. čes. 105: 982-984.
Kramer, W. (1966): Frontiers of neurological diagnosis in acquired toxoplasmosis. Psychiatr. Neurol. Neurochir. 69: 43-64.
Kretschmer, W., Schmid, E.E. (1955): Komplizierte Psychose bei Toxoplasma-Encephalitis. Arch. Psychiat. Nervenkr. 193: 38-48.
Kvalsvig, J.D. (1981): The effects of Schistosomiasis haematobium on the activity of school children. S. Afr. Med. J. 60: 61-64.
Kvalsvig, J.D. (1988): The effects of schistosomiasis on spontaneous play activity in black schoolchildren in the endemic areas. An ethological study. Psychoneuroendocrinology 23: 733-739.
Ladee, G.A., Scholten, J.M., Posthumus Meyes, F.E. (1966): Diagnostic problems in psychiatry with regard to acquired toxoplasmosis. Psychiatr. Neurol. Neurochir. 69: 65-82.
Lepš, J. (1996): Biostatistika. Jihočeská univerzita, České Budějovice.
Levav, M., Mirsky, A.F., Schantz, P.M., Castro, S., Cruz, M.E. (1995): Parasitic infection in malnourished school children: effect on behavior and EEG. Parasitology 110;103-111.
Levine, N.D. (1992): The protozoan phylum Apicomplexa. Volume 2. CRC Press Inc., Boca Raton, Florida.
Mackie, M J., Fiscus, A.G., Pallister, P. (1971): A study to determine causal relationships of toxoplasmosis to mental retardation. Am. J. Epidem. 94: 215-221.
Minto, A., Roberts, F.J. (1959): The psychiatric complications of toxoplasmosis. The Lancet 275: 1180-1182.
Monjour, L., Niel, G., Palminteri, R., Sidatt, M. Daniel-Ribeiro, C., Alfred, C., Gentlini, M. (1983): An epidemiological survey of toxoplasmosis in Mauritania. Trop. geogr. Med. 35: 21-25.
Moore, J., Gotelli, N.J. (1990): Phylogenetic perspective on the evolution of altered host behaviours: a critical look at the manipulation hypothesis. In: Barnard, C.J., Behnke J.M. (eds.) Parasitism and host behaviour. Taylor and Francis, London, New York, Philadelphia, 193-229.
Moore, J (1995): The behavior of parasitized animals. BioScience 45: 89-96.
Morell, V. (1997): How the malaria parasite manipulates its hosts. Science. 278: 223.
Nelson, G.S. (1990): Human behaviour and the epidemiology of helminth infections: cultural practices and microepidemiology. In: Barnard, C.J., Behnke J.M. (eds.) Parasitsm and host behaviour. Taylor and Francis, London, New York, Philadelphia, 234-263.
Nemec, R., Nečas, S., Holková, R., Čatár, F., Porubenová, A. (1988): Titre toxoplasmových protilátok u pracovníkov s potenciálne vyššou možnosťou toxoplazmovej nákazy. Bratisl. Lek. Listy 89: 779-783.
Nokes, C., Grantham-McGregor, S.M., Sawyer, AW., Cooper, E.S., Robinson, B.A., Bundy, D.A.P. (1992): Moderate to heavy infection of Trichiuris trichiura affect cognitive function in Jamaican children. Parasitology 104: 539-547.
Nokes, C., Bundy, D.A.P. (1994): Does Helmith Infection Affect Mental Processing and Educational Achievement? Parasit. Today 10: 15-18.
O’Connell, S., Guy, E.C., Dawson, S.J., Francis, J.M., Joynson, D.H.M. (1993): Chronic active toxoplasmosis in an imunocompetent patient. 27: 305-310.
Owen, M.R., Trees, A.J. (1998): Vertical transmission of Toxoplasma gondii from chronically infected house (Mus musculus) and field (Apodemus sylvaticus) mice determined by polymerase chain reaction. Parasitology 116: 299-304.
Pariser, S.F., Zunich, J., Pinta, E.R. (1978): Toxoplasmosis masquerading as a psychotropic side effect. J. Clinic. Psychiatry. 39: 631-632.
Parker, M. (1992): Re-assessing disability: the impact of schistosomal infection on daily activities among women in Gezira Province, Sudan. Soc. Sc. Med. 35: 877-890.
Perea, E.J., Borobio V., (1974): Facteurs Épidemiologiques et Caractéres Cliniques de la Toxoplasmose en Espagne. Bull. Soc. Pathol. exot. 67: 183-193.
Petrovický, O., Kvapilík, J. (1955): Zkušenosti s toxoplasminovým testem ve Státní psychiatrické léčebně v Kroměříži na základě allergologických studií. Čas. Lék. čes. 94: 933-937.
Petrovický, O., Vojtěchovský, M. (1955): Klinický obraz získané toxoplasmosy s neurologického a psychiatrického hlediska. Čas. Lék. čes. 94: 930.
Poulin, R. (1995): „Adaptive“ changes in the behaviour of parasitized animals: A critical review. Inter. J. Parasitol. 25: 1371-1383.
Quilici, M., Ranque, P., Tounkara, A., Rougemont, A., (1976): La Toxoplasmose en République du Mali Approche épidémiologique. Acta Trop. 33: 229-239.
Rau, M.E. (1983): The open field behaviour of mice infected with Trichinella spiralis. Parasitology 86: 311-314.
Rau, M.E., Putter, L. (1984): Running responses of Trichinella spiralis-infected CD-1 mice. Parasitology 89: 579-583.
Ricciardi, I.D., Sabroza, P.C., Sandoval, E.D., Mayrink, W. (1978): Seropepidemiological study on the prevalence of human toxoplasmosis in Brazil. Rev. Microbiol. 9: 181-187.
Remington, J.S., Efron, B., Cavanaugh, E., Simon, H.J., Trejos, A. (1970): Studies on toxoplasmosis as measured by the Sabin-Feldman dye test. Trans. Royal Soc. Trop. Med. Hyg. 64: 252-267.
Remington, J.S. (1974): Toxoplasmosis in the adult. Bull. N. Y. Acad. Med. 50: 211-227.
Remington, J.S., Krahenbuhl, J.L. (1982): Immunology of Toxoplasma gondii. In: Nahmias, A.J., O’Reilly, R.J. (eds.): Immunology of human infection, Part 2. Plenum Publishing Corporation, New York. 327-371.
Resano, P.F., Pascoe, L.D., Zuniga, T.V. (1985): Encuesta a Sero-epidemiologica de Anticuerpos Anti-toxoplasma en la Republica Mexicana. Rev. Mex. Patol. Clin. 32: 8-20.
Riegrová, J., Ulbrichová, M. (1993): Aplikace fyzické antropologie v tělesné výchově a sportu (příručka funkční antropologie). Univerzita Palackého, Olomouc.
Robertson, J.S. (1965): Toxoplasma skin- and dye-test surveys of severely subnormal patients in Linconshire. J. Hyg. 63: 89-98.
Robinson, R.A. (1973): The potential role of cats in the epidemiology of Toxoplasma gondii infections. New Zealand Med. J. 77: 97-101.
Roizen, N., Swischer, Ch.N., Stein, MA., Hopkins, J., Boyer, K.M., Holfels, E., Mets, M.B., Stein, L., Patel, D., Meier, P., Withers, S., Remington, J., Mack, D., Heydemann, P.T., Patton, D., McLeod, R. (1995): Neurological and developmental outcome in treated congenital toxoplasmosis. Pediatrics 95: 11-20.
Říčan, P. (1975): Šestnáctifaktorový dotazník 16PF (příručka), 2. přepracované a rozšířené vydání. Psychodiagnostika, Praha.
Saathoff, M., Dogba, C., (1974): Sero-epidemiologische Untersuchungen zum Vorkommen von Toxoplasma-Infektionen bei Jugendlichen und Erwachsenen in Sud-Togo. Z. Tropenmed. Parasit. 25: 401-404.
Sacks, J.J., Delgado, D.G., Lobel, H.O., Parker, R.L. (1983): Toxoplasmosis infection associated with eating undercooked venison. Am. J. Epidemiol. 118: 832-838.
Savory, M.A. (1992): Selective effects of colds and influenza on human performance efficiency: A critical appraisal. Neuropsychobiology. 25: 153-160.
Sharma, S.P., Dubey, J.P. (1981): Quantitative survival of Toxoplasma gondii tachyzoites and bradyzoites in pepsin and in trypsin solution. Am. J. Vet. Res. 42: 128-130.
Schassan, H.H., Kaskara, S. (1971): Die Durchseuchung mit Toxoplasma gondii in Hamburg. Tropenmed. Parasit. 22: 167-177.
Schnurrenberger, P.R., Tjalma, R.A., Wentworth, F.H., Wentworth, B.B. (1964): An association of human reaction to intradermal toxoplasmin with degree of animal contact and rural residence. Am. J. Trop. Med. Hyg. 13: 281-286.
Simeon, D., Callender, J., Wong, M., Grantham-McGregor, S., Ramdath, D.D. (1994): School performance, nutritional status and trichiuriasis in Jamaican schoolchildren. Acta Pediatr. 83: 1188-1193.
Simeon, D.T., Grantham-McGregor, S.M., Callender, J.E., Wong, M.S. (1995): Treatment of Trichuris trichiura infection improves growth, spelling scores and school attendance in some children. J. Nutr. 125: 1875-1883.
Smith, A.P., Tyrrell, D.A.J., Al-Nakib, W., Barrow, P.G., Higgins, P.G., Leekam, S., Tricket, S. (1989): Effects and after-effects of the common cold and influenza on human performance. Neuropsychobiology 21: 90-93.
Smith Trail, D.R. (1980): Behavioral interactions between parasites and hosts: host suicide and the evolution of complex life cycles. Am. Naturalist 116: 77-91.
Solomons, N.W. (1993): Pathways to impairment of human nutritional status by gastrointestinal pathogens. Parasitology 107:S19-S30.
Stagno,S., Thiermann,E. (1973): Acquisition of toxoplasma infection by children in a developing country. Bull. Wld. Hlth. Org. 49: 627-631.
Stibbs,H.H. (1985): Changes in brain concentrations of catecholamines and indoleamines in Toxoplasma gondii infected mice. Ann. Trop. Med. Par. 79: 153-157.
Ševkunova, E.A., Melnikov, V.D., Džanpoladova, V.P., Gračeva, L.I., Carueva, T.V., Kerimova, Z.M. (1976): Sravnitelnaja očenka roli domašnich košek i sobak v epidemiologii toksoplazmoza. Ž. Epidemiol. 5: 64-68
Thalhammer, O. (1962): Congenital toxoplasmosis. The Lancet 278: 23-24.
Vijayamma, T., Sinniah, B., Yap, P.L. (1980): Toxoplasma gondii antibodies in Malaysians. Southeast Asian J. Trop. Med. Pub. Hlth. 11;119-125.
Wallace, G.D., Zigas, V., Gajdusek, C.D. (1974): Toxoplasmosis and cats in New Guinea. Am. J. Trop. Med. Hyg. 23: 8-14.
Warren, J., Sabin, A.B. (1942): The complement fixation reaction in toxoplasmic infection. Proc. Soc. Exper. Biol. Med. 51: 11-16.
Webster, J.P. (1994): The effect of Toxoplasma gondii and other parasites on activity levels in wild and hybrid Rattus norvegicus. Parasitology. 109: 583-589.
Webster, J.P., Brunton, C.F.A., Macdonald, D.W. (1994): Effect of Toxoplasma gondii upon neophobic behaviour in wild brown rats, Rattus norvegicus. Parasitology. 109: 37-43.
Yilmaz, S.M., Hopkins, S.H. (1972): Effects of different conditions on duration of infectivity of Toxoplasma gondii oocyst. J. Parasit. 58: 938-939.
Zástěra, M., Hubner, J., Pokorný, J., Seeman, J., Peychl, L., Kotrlík, J., Kubásek, M., Koutský, J. (1966): K otázce zdrojů lidské toxoplasmosy. Čs. Epidem. Mikrobiol. Imunol. 15: 340-345.
Zitková, Š. (1996): Změny v chování myši způsobené parazitickým prvokem Toxoplasma gondii. Diplomová práce. PřF UK Praha.